XiaoMi-AI文件搜索系统
World File Search System友永真

具有双果冻的真核DNA病毒的自然史
门前病毒(Kingdom Bamfordvirae,Realm varidnaviria)是多种病毒的广泛组合,其相对较短的双链DNA基因组(<50 kbp)产生了由双果冻 - 双果冻 - 卷胶卷蛋白构建的二十os虫。前肿瘤动物感染所有细胞结构域的宿主,证明其古老的起源,尤其是与真核生物的七个超级组中的六个有关。前肿瘤分子包括四个主要的病毒组,即Polinton,Polinton,例如病毒(PLV),病毒噬细胞和腺毒。我们使用蛋白质结构建模和分析来表明蛋白质的DNA聚合酶(PPOLBS),polins,病毒噬细胞和细胞质线性质粒涵盖了n-终末结构域与末端蛋白(TPS)的N-末端域同源物(TPS),例如原始prd1-涉及tpectiricotic andototic artectirIdotics和eukaryotic artirIdotics artirIdotic artirIdotic artineciridotics anden tectirifiridotic toNERIFIRIDICRIDOTICSIRIATICS -ETENIRIDOTIOTICTIRIDOTOCTIOTICTIRIDS复制启动,以病毒卵巢肿瘤 - 类半胱氨酸去泛素酶(votu)结构域为由。投票域可能是导致TP从大型PPOLB多肽裂解的原因,并且在腺毒中被灭活,其中TP是单独的蛋白质。许多PLV和转囊编码了Polinton的独特衍生物 - 例如保留TP,Fotu和PPOLB聚合棕榈域的PPOLB,但缺乏外核酸酶域,而含有一个超家族1个旋转酶结构域。分析了在真核前肿瘤前胞菌中,对投票域的存在/不存在和将PPOLB用其他DNA聚合酶代替,使我们能够概述其起源和进化的完整情况。

新西兰排放交易体系 (ETS) NZU 盈余建议
不得因报告内容或向第三方提供报告而对安永提出任何索赔或要求或任何诉讼或程序。安永将永远免于承担任何此类索赔、要求、诉讼或程序。在编制本报告时,安永已考虑并依赖客户和参与流程的其他利益相关者以及其他被认为可靠和准确的来源向我们提供的信息。安永并未被告知任何提供给它的信息或从公共来源获得的信息是虚假的,或任何重大信息被隐瞒。安永并不暗示,也不应被解释为安永已对提供给我们的任何信息执行了审计、验证或尽职调查程序。安永未独立验证任何此类信息,也不承担独立验证此类信息的任何责任或义务,也不对信息的准确性或完整性作出任何陈述。安永或其任何成员或员工均不承担任何责任,对于因向安永提供不正确信息而导致本报告中的错误而给任何人造成的任何损失或损害,亦不承担任何责任。

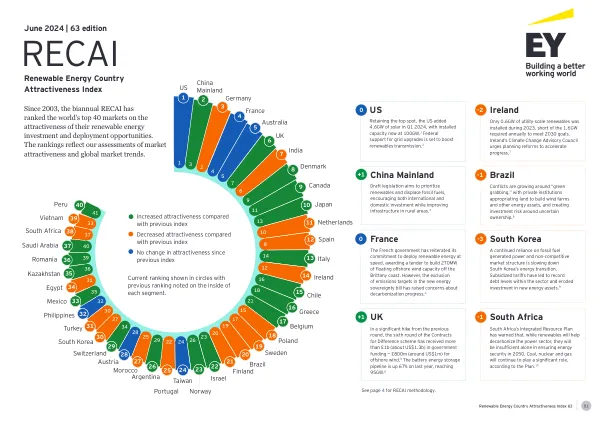
RECAI 63 前40名排名
安永是安永全球有限公司的全球组织,也可能指其一个或多个成员机构,每个成员机构都是独立的法人实体。安永全球有限公司是一家英国担保有限公司,不向客户提供服务。有关安永如何收集和使用个人数据以及个人根据数据保护法所享有的权利的描述,请访问 ey.com/privacy。安永成员机构不在当地法律禁止的地区执业。如需了解有关我们组织的更多信息,请访问 ey.com。

真核SMC蛋白在每个DNA处诱导-0.6的扭曲...
真核生物携带三种类型的结构性维持(SMC)蛋白复合物,冷凝蛋白,粘着素和SMC5/6,它们是ATP依赖性运动蛋白,通过DNA环挤出重塑基因组。SMCS调制DNA超螺旋,但仍未完全了解如何实现这一目标。在这里,我们提出了一个单分子磁性镊子测定法,该测定法直接测量每个回路 - 分解步骤中单个SMC诱导的扭曲程度。我们证明,所有三个SMC复合物都将相同的较大的负扭曲(即,链接数变化δk k k k占-0.6在每个回路 - 排除步骤中)中的挤压循环,与步长大小无关。使用ATP-Hydrolsyssys突变体和不可用的ATP类似物,我们发现ATP结合是ATPase循环期间的扭曲诱导事件,它与产生力的环路 - 分解步骤相吻合。所有三种真核SMC蛋白诱导相同数量的扭曲表明这些SMC复合物中常见的DNA环境解开机制这一事实。

经济和社会影响报告
安永会计师事务所 (“EY”) 受微软加拿大公司 (“微软”) 委托开展一项社会和经济影响研究。在编制本文件 (“报告”) 时,安永依赖来自第三方来源、微软、协会、学术和研究机构以及公共部门的未经审计的数据和信息 (统称为“支持信息”)。如果我们在本报告发布后获得更多支持信息,安永保留修改本报告中提及的任何分析、观察或评论的权利。安永假定支持信息准确、完整且适用于本报告的目的。安永未审计或独立核实支持信息的准确性或完整性。因此,安永不对支持信息发表任何意见或其他形式的保证,也不对任何错误或遗漏或因任何人将本报告用于编制目的以外的任何目的而造成的任何损失或损害承担任何责任。

布里真德 2030 年净零碳战略
如今,全球气候紧急状态已得到前所未有的政治认可。2018 年 10 月,政府间气候变化专门委员会 (IPCC) 发布了一份报告,详细说明了我们应对大规模气候崩溃所剩无几的时间。他们表示,我们必须在 2030 年之前将碳排放量削减到将全球变暖限制在比工业化前水平高出 1.5°C 的水平。这最终促成了第 15 届联合国气候变化大会 (COP) 达成的《巴黎气候变化协定》,189 个国家联合批准了一项具有法律约束力的承诺,采取行动限制本世纪全球气温上升 1 ,而最近在 2021 年 10 月于格拉斯哥举行的第 26 届联合国气候变化大会上,各方商定了具体的气候变化缓解行动,以停止毁林、减少甲烷排放并停止煤炭排放。

精确可调的,可抑制的真核转录控制器以实现遗传发现
基因的抽象条件表达和表型的观察仍然是生物学发现的核心。当前方法可启用开/关或不确定的分级基因表达。,我们开发了一个“脾气好”的控制器WTC 846,用于精确可调,分级,生长条件在酿酒酵母中基因的独立表达。受控的基因是由核酸脑抑制的强烈半合成启动子表达的,这也抑制了其自身的合成。基础表达被第二秒消除,“零”阻遏物。自动层环降低细胞对细胞的变化,同时通过化学诱导剂对蛋白质表达进行精确调整。WTC 846 allelic strains in which the controller replaced the native promoters recapitulated known null phenotypes ( CDC42, TPI1 ), exhibited novel overexpression phenotypes ( IPL1 ), showed protein dosage-dependent growth rates and morphological phenotypes ( CDC28, TOR2, PMA1 and the hitherto uncharacterized PBR1 ), and enabled cell cycle同步(CDC20)。WTC 846定义了一个“表达夹”,可以通过实验者在细胞蛋白丰度范围内调整蛋白质剂量,而设定点周围的变化有限。

真核生物核糖体中药物结合残基的自然变异
标题 真核核糖体中药物结合残基的天然变异 作者 Lewis I. Chan 1,& 、Chinenye L. Ekemezie 1,& 、Karla Helena-Bueno 1 、Charlotte R. Brown 1 、Tom A. Williams 2,* Sergey V. Melnikov 1,* 附属机构 1 纽卡斯尔大学生物科学研究所,英国泰恩河畔纽卡斯尔,NE2 4HH 2 布里斯托尔大学生物科学学院,英国布里斯托尔,BS8 1TQ & 贡献相同 通讯 * 通讯地址:tom.a.williams@bristol.ac.uk 和 sergey.melnikov@newcastle.ac.uk 摘要 针对真核核糖体的药物作为研究工具和针对癌症、真菌和其他致病性的潜在疗法正变得越来越重要真核生物。然而,由于缺乏比较研究,我们目前不知道有多少真核生物拥有与人类相同的核糖体药物结合位点,以及有多少与人类有显著差异。目前,这种知识上的差距因真核生物基因组中存在假基因而加剧,由于我们无法区分真正的突变、假基因和测序伪影,使得这些比较分析具有挑战性。在本研究中,我们通过使用一种利用物种间进化关系的新方法解决了这个问题。使用这种方法,我们确定了 8,563 种代表性真核生物中 58 种核糖体药物结合残基的序列变体,追溯了这些变异的进化历史,从 20 亿年前真核生物的出现到它们随后分化成不同的谱系。出乎意料的是,我们发现酵母和人类(通常用作研究核糖体/药物相互作用的模型真核生物)与大多数其他真核生物不同,因为 rRNA 替换主要发生在动物和真菌中,但在大多数其他真核生物中不存在。此外,我们证明了以前在常见病原体利什曼原虫和疟原虫中发现的结构变异,这些变异被视为少数真核生物物种所特有的,但实际上为大量真核生物所共有。值得注意的是,一些真核生物谱系的核糖体药物结合位点与人类的差异比人类与细菌的差异更大。总体而言,我们的研究提供了真核生物核糖体药物结合位点进化的最完整概述(在单个物种、单个残基和单个药物的水平上),确定了与人类相比具有结构不同的核糖体药物结合位点的真核生物谱系。这些发现为利用核糖体靶向药物作为研究工具和开发针对真核寄生虫的谱系特异性抑制剂开辟了新的途径。