XiaoMi-AI文件搜索系统
World File Search System受扰

植物防御信号通路之间的串扰
植物暴露于非常不同的攻击者,包括微生物病原体和草食昆虫。为了保护自己,植物已经发展了防御策略,以抵消潜在的入侵者。植物防御信号研究的最新进展表明,根据遇到的入侵者的类型,植物能够差异激活诱导,广谱防御机制。植物激素水杨酸(SA),茉莉酸(JA)和乙烯(ET)是防御信号通路网络中的主要参与者。在SA-,JA-和ET依赖性信号通路之间的串扰被认为与对防御反应进行微调有关,最终导致了防御反应的最佳组合以抵抗入侵者。这些信号化合物的生物合成途径的基因工程以及模仿其作用方式的保护化学物质的开发为开发新策略的作物保护提供了有用的工具。但是,有证据表明,对微生物病原体的抗药性与对草食昆虫的抗药性之间的抗性:一旦植物的条件表达对微生物病原体的抗性,它可能会更容易受到食草动物的攻击,而反之亦然。然而,病原体和抗昆虫抗性之间的贸易证据是矛盾的。本综述集中于有关SA-,JA-和ET依赖性诱导对微生物病原体和草食性昆虫的抗性的最新实验证据。此外,我们将解决以下问题,无论是通过基因工程或通过使用防御信号的植物保护剂来操纵国防信号通路,是否会增强植物对潜在入侵者的免疫力,还是将成为作物保护策略的负担。

Toll样受体和整联蛋白串扰
免疫系统识别病原体和抗原水平的入侵微生物。Toll样受体(TLR)在针对病原体的第一线防御中起关键作用。TLR的主要功能包括细胞因子和趋化因子的产生。TLR与其他受体共享常见的下游信号通路。围绕TLR旋转的串扰相当复杂而复杂,强调了免疫系统的复杂性。通过TLRS产生的细胞因子和趋化因子的蛋白鱼可能会受其他受体的影响。整合素是在许多不同细胞上表达的关键异二聚体粘附分子。有一些研究描述了TLR和整联蛋白之间的协同或抑制性相互作用。因此,我们回顾了TLR和整合素之间的串扰。了解串扰的性质可以使我们能够通过整合素来调节TLR功能。
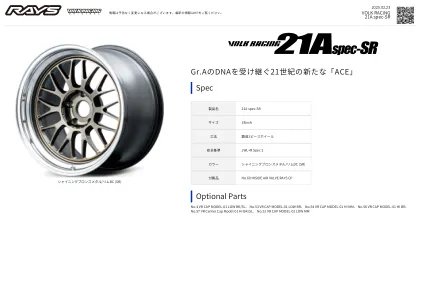
21世纪的新“ ACE”,继承了Gr.A的DNA规格...
尺寸图补充[Caliper清除率]▼:Impreza(GDB 4Pot)▽:Impreza(GDB&GRB 4POT)θ:WRX STI(4pot)η:WRX STI(6pot)§:GR86/BRZ BREMBO(GR86/BRZ BREMBO)( :rc-f#:silvia(S15)△:天际线(R34GT-R,CPV35,CKV36, HV37,RV37/400R)▲:天际线(R34 GT-R)□:Skyline(CKV36,HV37,RV37/400R)α:Skyline(CKV36,HV37) ●:Fairlady Z(Z34,RZ34)ω:Fairlady Z(Z33)☆:Lancer Evo(VII-IX,X)★:Lancer Evo.x ∑:Lancer Evo(vii-ix)◆:GT-R(R35/φ380转子)◇:GT-R(R35/φ380,φ390rotor)〓:GR Yaris(rz)ψ:Roadster(nd)/124 Spider Brembo * spider Brembo * Caliper Compatible Veyals的示例仅供参考。请咨询零售商,然后选择要安装的尺寸。

超越边界:对机器人技术的接受与抵制
例如,当非残疾人士利用机器人技术来增强他们的身体或智力能力并更好地在社会中生活时,他们可能不会感到自己被物化。但是,考虑到机器人技术所带来的超能力是外界赋予的机械能力或者物质能力,因此也可以理解为人是自愿物化的。 当我们考虑这种情况时,人类是否会抵制无休止地卷入对物质能力的竞争以及人类价值重心向物质能力转移的趋势?如果跨性别者或新人类的存在成为永久特征,那么基于精英统治或经济实力的社会差距是否会扩大?相反,如果建立一个基于社会共识的体系,让任何人都可以成为超人类,获得一定的身体和智力能力,甚至鼓励这种做法,是否会带来一个更加平等和包容的社会,社会差距是否会消失?当这一切发生时,成为一个符合自己性格的超人类,是否会成为一种可以被轻易接受和适应的事情,就像在虚拟空间中为自己的角色收集配饰的感觉一样?

神经网络的非微扰重正化-...
在最近的一项工作 [ 1 ] 中,Halverson、Maiti 和 Stoner 提出了一种用威尔逊有效场论来描述神经网络的方法。无限宽度极限被映射到自由场论,而有限 N 个校正则由相互作用(作用中的非高斯项)考虑。在本文中,我们研究了这种对应的两个相关方面。首先,我们在这种情况下评论了局部性和幂计数的概念。事实上,这些通常的时空概念可能不适用于神经网络(因为输入可以是任意的),然而,重正化群提供了局部性和缩放的自然概念。此外,我们还评论了几个微妙之处,例如数据分量可能不具有置换对称性:在这种情况下,我们认为随机张量场论可以提供自然的概括。其次,我们通过使用 Wetterich-Morris 方程提供非微扰重正化群的分析,改进了 [1] 中的微扰威尔逊重正化。与通常的非微扰 RG 分析的一个重要区别是,只知道有效 (IR) 2 点函数,这需要谨慎设定问题。我们的目标是提供一种有用的形式化方法,以非微扰方式研究超越大宽度极限(即远离高斯极限)的神经网络行为。我们分析的一个主要结果是,改变神经网络权重分布的标准差可以解释为网络空间中的重正化流。我们专注于平移不变核并提供初步的数值结果。

利用量子加扰改进计量学 - NSF-PAR
量子扰乱描述了信息在量子系统中扩散到许多自由度的过程,这样信息就不再是本地可访问的,而是分布在整个系统中。这个想法可以解释量子系统如何变成经典系统并获得有限的温度,或者在黑洞中,物质落入的信息是如何被抹去的。我们探测了相空间中双稳态点附近的多粒子系统的指数扰乱,并将其用于纠缠增强计量。时间反转协议用于观察计量增益和不按时间顺序的相关器同时呈指数增长,从而通过实验验证了量子计量和量子信息扰乱之间的关系。我们的结果表明,能够以指数速度快速产生纠缠的快速扰乱动力学对实际计量很有用,可产生超出标准量子极限 6.8(4) 分贝的增益。E

B7/CD28 和 EGFR 通路之间的串扰
摘要表皮生长因子受体 (EGFR) 中的体细胞激活突变是癌症(例如非小细胞肺癌 (NSCLC)、转移性结直肠癌、胶质母细胞瘤、头颈癌、胰腺癌和乳腺癌)中最常见的致癌驱动因素之一。针对 EGFR 信号通路的分子靶向药物已显示出强大的临床疗效,但患者不可避免地会出现获得性耐药。尽管针对 PD-1/PD-L1 的免疫检查点抑制剂 (ICI) 在多种癌症类型的部分患者中表现出持久的抗肿瘤反应,但它们对含有 EGFR 激活基因变异的癌症的疗效有限。越来越多的研究表明,新的 B7/CD28 家族成员(如 B7-H3、B7x 和 HHLA2)的上调与 EGFR 信号传导有关,并可能通过创建免疫抑制肿瘤微环境 (TME) 导致对 EGFR 靶向疗法的耐药性。在本综述中,我们讨论了 EGFR 信号传导对 PD-1/PD-L1 通路和新的 B7/CD28 家族成员通路的调节作用。了解这些相互作用可能有助于制定联合治疗策略,并可能克服当前对 EGFR 靶向疗法的耐药性挑战。我们还总结了抗 PD-1/PD-L1 疗法在 EGFR 突变癌症中的临床数据,以及

皮肤菌群与免疫系统之间的串扰
皮肤微生物组由多样化的微生物及其相关产品组成。这些微生物直接与宿主细胞相互作用,并受到皮肤免疫反应和外部因素(例如抗生素)的影响。皮肤微生物组的好处包括在早期生命中建立免疫学耐受性,抗菌药物的产生和免疫调节的代谢产物,促进伤口愈合,增强屏障功能以及迁移,代谢性,代谢和皮肤细胞功能的调节。相比之下,皮肤微生物组中的病原体和病原体会引起疾病,并与皮肤疾病有关(图1)。皮肤微生物组和宿主之间的串扰非常复杂,并且仍然存在许多知识差距。了解管理皮肤微生物生态的“规则”及其失调对宿主免疫的影响将是推进这一领域并意识到使用微生物及其代谢物用于治疗目的的希望的关键。

骨与脑之间的分子和细胞串扰
分子法技术,包括蛋白质组学,已使关键信号通路阐明了介导大脑和骨组织之间双向通信的关键信号通路。在这里,我们简要摘要研究了研究跨组织细胞通信的骨 - 脑轴的需求。明确的临床和分子证据表明骨骼和脑细胞之间的生物学相互作用和相似性。在这里,我们回顾了目前研究大脑和骨骼疾病的质谱技术,分别重点是神经退行性疾病和骨关节炎/骨质疏松症。在分子水平上进一步研究了蛋白质,神经肽,骨化剂和激素在与骨骼和脑部疾病相关的分子途径中的作用是至关重要的。使用质谱和其他OMIC技术来分析这些跨组织信号传导事件和相互作用将有助于我们更好地了解疾病的进展和合并症,并有可能确定治疗性干预措施的新途径和目标。蛋白质组学测量值特别有利于提取信号传导,分泌和循环分析物的作用,并识别与年龄相关疾病有关的含量和代谢途径。

量子信息处理器的串扰理论及其缓解方法
本论文的第二部分详细介绍了我们通过实验表征和有效缓解固定频率超导量子比特串扰的尝试。我们遇到的第一个障碍是了解串扰对系统的影响。当串扰较弱时,现有方法很难奏效,因此我们开发了一种测量串扰的新方法。我们需要解决的第二个问题是验证我们的模型是否正确。利用我们第一次测量的结果,我们将预测的演变与与测量过程截然不同的环境中实验数据进行比较。我们发现实验和理论之间具有很好的一致性,表明该模型是合理的。本次调查中最后一个未解决的难题是使用此模型来缓解串扰,我们的研究仍在进行中。