XiaoMi-AI文件搜索系统
World File Search System外源


飞机润滑油和其他 10 种物品
5.有关保证金事项 (1)有关投标保证金或合同保证金事项:免收 (2)中标人不签订合同的,将收取中标金额5/100的罚款。 6. 无效投标 (1)不具备本公告所列参与投标所需资格的人员投标以及违反投标条件的人员投标 (2)未收到规格说明书的人员投标 (3)不按照投标指南进行投标和签订合同的人员 地点:小松岛空军基地,航空中队会议室 7. 是否需要准备合同 必须立即准备合同。但合同金额不超过150万日元,且无特殊条款的情况下,可以开具收据。 8 适用合同条款 买卖合同的一般条款 9 其他 (1)收到规范书时,应当提交资格审查结果通知书复印件。 (2)目前中止招标的企业原则上不允许进行分包。但有关部会暂停提名权认定有确实不可避免的事由时,不在此限。 (3)投标联系方式:1. 〒773-8601 德岛县小松岛市和田岛町须田4-3

Hangzhou Jiuyuan Gene Engineering Co.,Ltd。杭州九源... 2023环境社会和治理报告 AIM疫苗有限公司AIM疫苗有限公司艾美疫苗股份有限公司 年度的年度业绩公告... AIM疫苗有限公司AIM疫苗有限公司艾美疫苗股份有限公司 远大医药集团有限公司 电子商品控股有限公司
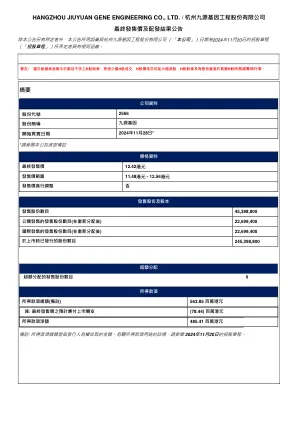
于临时股东大会日期,245,398,800股,包括,包括136,302,015股非上市股份及109,096,785股h股。于临时股东大会日期,(i)(i)本公司概无持有库存股份(包括持有或存置于由香港中央结算有限公司建立及运作的中央结算及交收系统的任何库存股份),因此于临时股东大会上并无库因此于临时股东大会上并无库(I),因此于临时股东大会上并无库因此于临时股东大会上并无库(ii)(ii)本公司概无购回待注销之股份。出席临时股东大140,027,027,52525252525252525252525252525252525252525252525252525252525252525.0.0.0.06%6%6%。

Hangzhou Jiuyuan Gene Engineering Co.,Ltd。杭州九源...Hangzhou Jiuyuan Gene Engineering Co.,Ltd。杭州九源...
在EGM之日,有权参加EGM提议的决议的股份和股份总数为245,398,800,其中包括136,302,015股份,其中包括136,302,015股股份和109,096,096,785小时。在EGM之日起,(i)公司没有持有的财政部股份(包括由香港证券清算公司建立和运营的中央清算和和解系统持有或存放的任何财政部股票),因此没有在EGM上行使财政部股票的投票权; (ii)没有公司回购待定的股票。股东的数量和授权的代理人参加了EGM。股东和授权的代理人持有140,027,525股股票,占EGM,约占具有投票权的已发行股票总数的57.06%。

Hangzhou Jiuyuan Gene Engineering Co.,Ltd。杭州九源... CSPC制药集团有限公司 年度报告
(4)股东须以书面形式委任代表,代表委任表格由股东或由其以书面正式授权的代理人签,代表委任表格应加盖公司印章或由其董事或以书面形式正式授权的,代表委任表格应加盖公司印章或由其董事或以书面形式正式授权的,则授权书或其他授权文件必须经公24小时前(即不迟于2025年3月4上午九时正前(香港时间)((())(())将上述文件送达h股证券登记处香港中央证券登h股证券登记处香港中央证券登173号合和中心17m楼)

10、11号楼应急电源改造工程
2024年9月17日 - 8 文件程序。建筑工程所需的申请和文件必须按照政府规定的标准格式准备和提交。 9 保密。未经监理人员许可,不得泄露施工期间获得的任何信息。

电熨斗,100V1500W 及其他4个
2024年11月8日——国防部竞标资格。(各部委统一资格)。“商品制造。”“商品销售。”“服务... *除产品名称和标准要求外,同等或更高质量的产品(包括其他公司的产品)均被认可...

冈山大学资源植物科学研究所报告
植物压力的研究核心科学大气压力单元植物光适应研究小组1组环境反应系统2功能性生物分子发现组组3土壤应力单位植物应力生理4植物分子生理学组分子生理学5生物应力单元组的植物 - 微生物相互作用6组植物 - 内部相互作用7植物免疫设计组8植物环境微生物学9大麦和野生植物资源中心遗传资源遗传资源单位遗传资源组基因组多样性10应用基因组学单位遗传资源和功能组11综合基因组育种12
00E250330826战争1..1
公众持股量,本公司已发⾏股本总额约,本公司已发⾏股本总额约28.44%将计算在公众持股量内,符合上市规则第,8.08条规定的最低百分比。,(i),紧随全球发售后,概无承配⼈将单独获配售本公司经扩⼤已发⾏股本10%以上;(ii)于上市后,除和达香港及杭⾦投,除和达香港及杭⾦投,共29,829,738 股股份(相当于我们的已发⾏股本总数约12.1616%)市时并无持有公众⼈⼠所持h股50%以上,符合上市规则第8.08(3)条及第8.24条的规定;及(iv)上市时至少有300名股东,符合上市8.08(2)8.08(2)条规定。条规定。开始买卖情况下,h股股票⽅会于香港时间2024年11⽉11⽉28⽇(星期四) (⽇(星期 ⽇(星期)上午九时正在联交所开始买卖。H股将以每手200股h股进⾏买卖。h股进⾏买卖。h股进⾏买卖。2566。