XiaoMi-AI文件搜索系统
World File Search System天冬


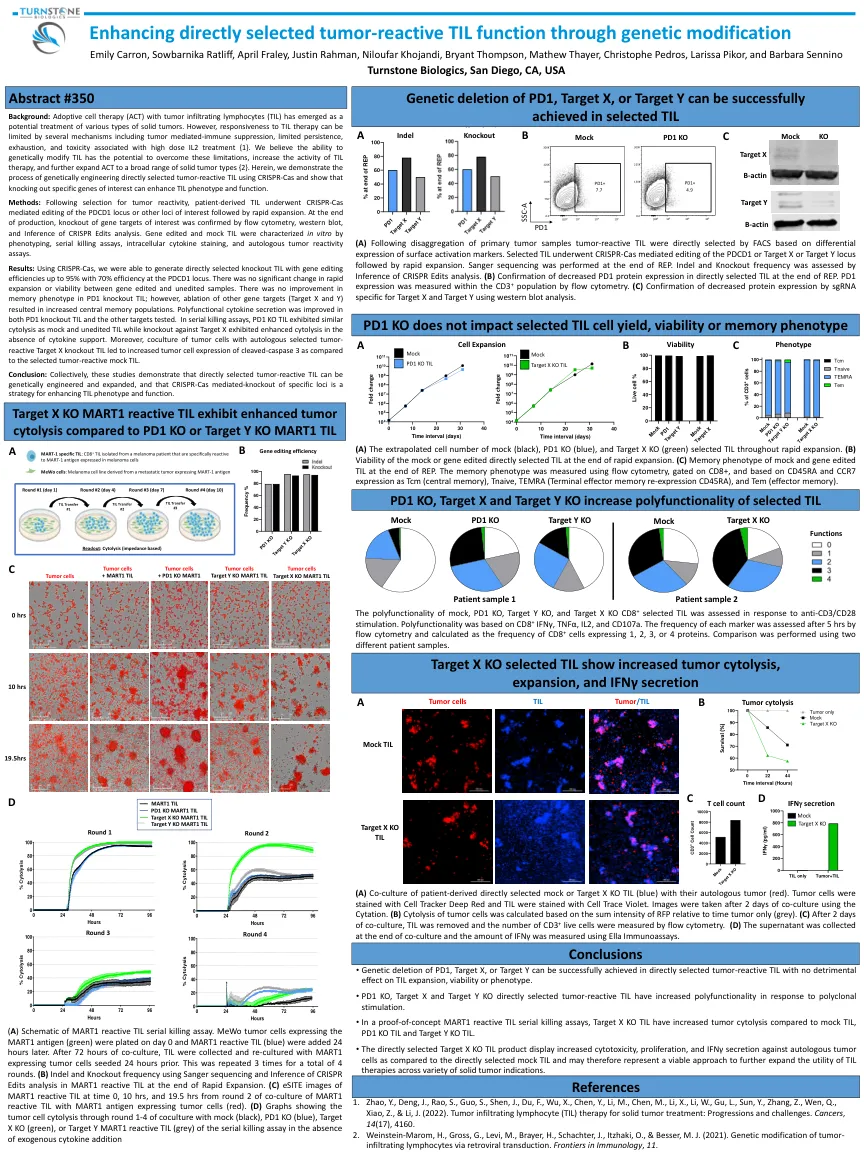
通过基因改造增强直接选择的肿瘤反应性 TIL 功能
结果:使用 CRISPR-Cas,我们能够生成直接选择的敲除 TIL,其基因编辑效率高达 95%,PDCD1 基因位点的效率为 70%。基因编辑和未编辑样本之间的快速扩增或活力没有显著变化。PD1 敲除 TIL 的记忆表型没有改善;然而,其他基因靶标(靶标 X 和 Y)的消融导致中枢记忆群体增加。PD1 敲除 TIL 和测试的其他靶标中的多功能细胞因子分泌均得到改善。在连续杀伤试验中,PD1 KO TIL 表现出与模拟和未编辑 TIL 相似的细胞溶解,而靶标 X 的敲除在没有细胞因子支持的情况下表现出增强的细胞溶解。此外,与选定的肿瘤反应性模拟 TIL 相比,肿瘤细胞与自体选定的肿瘤反应性靶标 X 敲除 TIL 共培养导致肿瘤细胞裂解半胱天冬酶 3 的表达增加。

醛介导的天冬酰胺生物合成抑制对糖尿病和酒精中毒的影响
在HCHO处理的LN-18细胞中进行了分析,揭示了天冬酰胺消耗的证据,尽管效果比HEK293T细胞弱(图S5†)。该观察结果至少部分是由于使用非二元胎牛血清的使用,该胎儿血清含有相对较高的天冬水平。与此相一致,与非二元胎儿牛血清一起孵育的HEK293T细胞未观察到氨基酸的耗竭(图S6和S7†)。半胱氨酸分别与HCHO和ACH反应,分别给出硫脯氨酸和2-甲基噻唑烷-4-羧酸(MTCA)(图1)。9,10,27用HCHO或ACH对HEK293T细胞的处理分别导致硫丙啉和MTCA水平升高(图1b)。在HCHO处理的LN-18细胞中也观察到了硫代丙烯的形成(图S5†)。在血28和人类寄生虫中报道了半胱氨酸和ACH对MTCA的反应; 29这里提供的证据还表明,MTCA可能发生在人类细胞中。半胱氨酸-MGO加合物不是

细胞凋亡与癌症关系的系统评价
细胞死亡途径最早由罗伯特·霍维茨在研究秀丽隐杆线虫等低等生物的细胞命运时发现,这最终帮助他获得了 2002 年诺贝尔生理学或医学奖。人们对细胞死亡机制有很多了解,包括从细胞内部和免疫系统 [1] 。其中最重要的一种机制是细胞凋亡,这是一种程序性细胞死亡,可有效清除受损细胞,例如在发育过程中或脱氧核糖核酸 (DNA) 损伤后 [2] 。细胞凋亡的一个重要特征是它从根本上通过一种叫做半胱天冬酶的丝氨酸蛋白酶亚型发挥作用,半胱天冬酶是一种半胱氨酰蛋白酶,可以通过蛋白水解裂解不同的细胞核和细胞质成分。这些胱天蛋白酶由11个成员组成,分为三大组,其中第二组(胱天蛋白酶2、3、7)和第三组(胱天蛋白酶6、8、9、10)参与细胞凋亡。胱天蛋白酶最终依靠不同的信号通路导致细胞的破坏[3]。细胞凋亡的发病机制复杂,涉及两种主要信号通路:外在和内在。两者都会激活效应凋亡胱天蛋白酶,最终导致细胞凋亡特征性的形态和生化改变[4,5]。决定细胞是否凋亡的最重要因素之一是促凋亡和抗凋亡蛋白调节剂之间的平衡。在癌前病变中,DNA损伤可诱导细胞凋亡以清除潜在的有害细胞,从而阻止肿瘤生长。相反,凋亡的紊乱会导致细胞增殖不受控制、癌症发展以及癌症对药物疗法的耐药性 [6]。癌细胞通常会过度表达不同的蛋白质,这些蛋白质在抵抗细胞凋亡的级联反应中起着重要作用。癌细胞诱导的多种机制将它们从程序性细胞死亡中拯救出来,尤其是通过抗凋亡分子的过度表达 [7]。事实上,大多数凋亡信号研究依赖于 B 细胞淋巴瘤 2 同源性 3 (BH3) 蛋白 [8]。促存活和促死亡的 BH3 蛋白之间存在平衡。当这种平衡偏向促死亡的 BH3 蛋白时,往往会发生细胞凋亡,但是当它偏向促存活蛋白时,就会导致存活信号的激活,从而导致癌症等病理状况。

细胞凋亡与癌症关系的系统评价
细胞死亡途径最早由罗伯特·霍维茨在研究秀丽隐杆线虫等低等生物的细胞命运时发现,这最终帮助他获得了 2002 年诺贝尔生理学或医学奖。人们对细胞死亡机制有很多了解,包括从细胞内部和免疫系统 [1] 。其中最重要的一种机制是细胞凋亡,这是一种程序性细胞死亡,可有效清除受损细胞,例如在发育过程中或脱氧核糖核酸 (DNA) 损伤后 [2] 。细胞凋亡的一个重要特征是它从根本上通过一种叫做半胱天冬酶的丝氨酸蛋白酶亚型发挥作用,半胱天冬酶是一种半胱氨酰蛋白酶,可以通过蛋白水解裂解不同的细胞核和细胞质成分。这些胱天蛋白酶由11个成员组成,分为三大组,其中第二组(胱天蛋白酶2、3、7)和第三组(胱天蛋白酶6、8、9、10)参与细胞凋亡。胱天蛋白酶最终依靠不同的信号通路导致细胞的破坏[3]。细胞凋亡的发病机制复杂,涉及两种主要信号通路:外在和内在。两者都会激活效应凋亡胱天蛋白酶,最终导致细胞凋亡特征性的形态和生化改变[4,5]。决定细胞是否凋亡的最重要因素之一是促凋亡和抗凋亡蛋白调节剂之间的平衡。在癌前病变中,DNA损伤可诱导细胞凋亡以清除潜在的有害细胞,从而阻止肿瘤生长。相反,凋亡的紊乱会导致细胞增殖不受控制、癌症发展以及癌症对药物疗法的耐药性 [6]。癌细胞通常会过度表达不同的蛋白质,这些蛋白质在抵抗细胞凋亡的级联反应中起着重要作用。癌细胞诱导的多种机制将它们从程序性细胞死亡中拯救出来,尤其是通过抗凋亡分子的过度表达 [7]。事实上,大多数凋亡信号研究依赖于 B 细胞淋巴瘤 2 同源性 3 (BH3) 蛋白 [8]。促存活和促死亡的 BH3 蛋白之间存在平衡。当这种平衡偏向促死亡的 BH3 蛋白时,往往会发生细胞凋亡,但是当它偏向促存活蛋白时,就会导致存活信号的激活,从而导致癌症等病理状况。

文章单一基于淀粉的水凝胶用于治疗性递送
摘要:水凝胶是各种治疗剂的输送系统的有趣材料,这主要是由于水湿网络和局部和持续的药物释放。在此,通过施加简便的合成并提议为新型的治疗分子递送系统而产生具有增强降解速率的单个基于淀粉的水凝胶。淀粉用钠周期氧化在水中和轻度条件下,以产生醛衍生物,在冻结过程后,允许淀粉衍生物紧凑和稳定的水凝胶。氧化淀粉还通过Schiff碱反应与天冬酸酯交联,以将活性分子直接连接到多糖结构。这些材料在结构和形态学上都是表征的,随着时间的推移,吸附和释放的能力通过QNMR光谱证明了活性分子。在Cal-27细胞系(口服鳞状细胞癌)上评估了细胞毒性。结果表明,由于细胞培养基的肿胀能力,合成的水凝胶导致细胞上的“冷冻增殖”状态。与未处理的对照相比,通过流式细胞仪数据表明,水凝胶在细胞中诱导的“早期凋亡”和更多的“晚期凋亡”。由于所提出的材料能够控制细胞的增殖,因此它们可以在精确治疗应用领域开放新情况。

caspase- 9抑制作用触发了基于HSP90的化学疗法
抽象背景抗塑性化学疗法在引起免疫原性死亡(ICD)时非常有效,从而诱导抗肿瘤免疫反应甚至消除肿瘤。然而,激活的胱天蛋白酶是大多数癌症化学治疗剂的标志,使凋亡在免疫学上保持沉默。它们是否对于化学疗法诱导的细胞死亡和体内细胞的凋亡清除率仍然难以捉摸。方法进行了基于理性细胞的抗癌药物库筛查,以探索在凋亡caspase抑制下的免疫原性凋亡途径和治疗靶标。基于这种筛选,caspase抑制在增强化学疗法诱导的抗肿瘤免疫力和作用机理方面的潜力通过各种细胞和小鼠模型研究了。结果热休克蛋白90(HSP90)抑制激活肿瘤细胞中的胱天蛋白酶,产生丰富的基因组和线粒体DNA片段,并导致细胞凋亡。同时,它劫持了caspase-9信号传导以抑制固有的DNA感应。Pharmacological blockade or genetic deletion of Caspase-9 causes tumor cells to secrete interferon (IFN)- β via tumor intrinsic mitochondrial DNA/the second messenger cyclic GMP–AMP (cGAS) /stimulator of interferon genes (STING) pathway without impairing Hsp90 inhibition-induced cell death.重要的是,CASPASE-9和HSP90抑制均可触发ICD,从而释放了许多损伤相关的分子模式,例如高摩动式组盒蛋白1,ATP和I型IFN和IFNS型IFN在体外和显着的抗肿瘤效应。此外,联合处理还通过上调编程的死亡配体1(PD-L1)来诱导适应性抗性。其他PD-L1阻滞可以进一步克服这种获得的免疫阻力并实现完全的肿瘤回归。结论caspase-9信号传导有选择地挑衅基于HSP90的化学疗法介导的肿瘤先天感应,从而导致CD8 + T细胞依赖性肿瘤控制。我们的发现暗示胱天冬酶途径的药理调节增加了化学疗法诱导的凋亡的肿瘤内在感应和免疫原性,

BmSPP 是家蚕的一种抗病毒基因
SPP 是一种 GXGD 型膜内裂解天冬氨酰蛋白酶,具有 9 个跨膜结构域,可裂解疏水脂质双层中的跨膜蛋白( 1 , 2 )。SPP 在整个进化过程中表现出高度的保守性,广泛存在于各种真核生物中,包括真菌、原生动物、植物和动物( 3 )。它具有广泛的生物学功能:通过消除前体信号肽酶 (SP) 裂解后在内质网 (ER) 中积累的信号肽来调节 ERAD 通路( 4 );与错误折叠的膜蛋白结合并形成参与体内自噬的大型寡聚复合物( 5 );通过水解信号肽来控制正常的免疫监视,促进表位片段的释放,保护细胞免受自然杀伤细胞 (NK) 的攻击 ( 6 );与病毒蛋白相互作用,影响病毒的加工和复制,或作为病毒逃避宿主免疫系统的手段 ( 4 , 7 – 9 )。敲低或抑制 SPP 会极大地影响生物体自身对病毒的抵抗力。SPP 介导的裂解负责将丙型肝炎病毒 (HCV) 核心蛋白引导到脂滴,这是病毒出芽和核衣壳组装的关键步骤。研究表明,使用抑制剂抑制 SPP 可以阻碍 HCV 增殖 ( 7 , 8 , 10 )。在感染过程中,单纯疱疹病毒 (HSV) 利用其糖蛋白 K (gK) 与 SPP 结合,促进 HSV-1 复制。SPP 诱导的敲除小鼠的病毒潜伏期显著缩短,使用 SPP 抑制剂后病毒复制也显著减少 ( 9 , 11 )。SPP 在猪瘟病毒 (CSFV) 核心蛋白的加工和成熟过程中起着重要作用,使用 (Z-LL) 2-酮抑制 SPP 可显著降低 CSFV 的活力 ( 12 )。这些实例凸显了 SPP 在病毒感染中的深远意义,表明针对宿主 SPP 可能是一种非常有效的抗病毒策略。家蚕(Bombyx mori)因其独特的吐丝特性而成为一种经济昆虫。然而,家蚕生产经常受到各种蚕业疾病的困扰。在这些疾病中,BmNPV 是最严重和最昂贵的病毒性疾病,导致严重的蚕业损失。考虑到 SPP 的特性,我们研究了编辑 BmSPP 是否可以提高家蚕对 BmNPV 的抵抗力。我们的预期是编辑 BmSPP 会产生抗性菌株。NPV 是一种存在于多种节肢动物中的杆状病毒,可感染 8 个目 600 多种昆虫,包括鳞翅目、膜翅目、双翅目、鞘翅目等(13)。它是一种具有双链环状 DNA 基因组的 DNA 病毒,因其基因组被包裹在杆状核衣壳中而得名(14)。BmNPV 在感染过程中产生两种类型的病毒颗粒:包涵体衍生病毒 (ODV) 和芽生病毒 (BV)。杆状病毒对宿主幼虫的感染是由 ODV 引起的,随后,BV 导致宿主的全身感染(15)。杆状病毒经口腔进入宿主,经前肠进入中肠,在中肠碱性环境中释放ODV。然后ODV直接与中肠细胞膜融合,释放核衣壳进入细胞质,导致原发性感染(14)。在宿主体内,病毒利用宿主自身的环境在宿主细胞内复制

铁凋亡和杯凋亡
缩写:5-FU,5-氟尿嘧啶;AA-CoA,花生四烯酸辅酶 A;ABCC1,ATP 结合盒,C 亚家族(CFTR/MRP),成员 1;ACC,无定形碳酸钙;ACLS4,酰基辅酶 A 合成酶家族 4;AdA-CoA,肾上腺酸辅酶 A;ALDH,醛脱氢酶;AML,急性髓细胞白血病;APC,抗原处理细胞;ARE,抗氧化反应元件;ART,青蒿素;BAX,BCL-2 相关 X 蛋白;BCL-2,B 细胞淋巴瘤 2;BTIC,脑肿瘤起始细胞;CBR,临床受益率;CLL,慢性淋巴细胞白血病;CNSI-Fe(II),碳纳米颗粒负载铁;CQ,氯喹;CRPC,去势抵抗性前列腺癌; CSC,癌症干细胞;CTL,细胞毒性 T 淋巴细胞;CuET,二乙基二硫代氨基甲酸铜 (II);DAMP,损伤相关分子模式;DFO,去铁胺;DHA,双氢青蒿素;DLAT,丙酮酸二氢硫酰赖氨酸残基乙酰转移酶成分;DMT1,二价金属转运蛋白 1;DOX,阿霉素;DRD2,多巴胺 D2 受体;DSF,双硫仑;EGFR,表皮生长因子受体;EMT,上皮-间质转化;ER,内质网;ETO,依托泊苷;FDX1,铁氧还蛋白 1;FER-1,铁抑制蛋白 1;FMN,基于框架的纳米剂;FPN1,铁转运蛋白 1;FTH1,铁蛋白重链 1; FTL1,铁蛋白轻链 1;GPX4,谷胱甘肽过氧化物酶 4;GSH,谷胱甘肽;GSS,谷胱甘肽合成酶;H 2 O 2,过氧化氢;HNC,头颈癌;HO-1,血红素加氧酶-1;ICD,免疫细胞死亡;ICIs,免疫检查点抑制剂;IDH1,异柠檬酸脱氢酶 1;IFN-γ,干扰素-γ;IREB2,铁反应元件结合蛋白 2;IREs,铁反应元件;IRP-2,铁调节蛋白 2;IRPs,铁调节蛋白;JAK,Janus 酪氨酸激酶;KEAP1,kelch 样 ECH 相关蛋白 1;KRAS,Kirsten 大鼠肉瘤病毒致癌基因同源物;LA,硫辛酸; LC3II,微管相关蛋白 1 轻链 3α;LDH,乳酸脱氢酶;LiMOFs,锂基金属有机骨架;LIPRO-1,利普司他丁 1;LOX,脂氧合酶;LPCAT3,溶血磷脂酰胆碱酰基转移酶 3;MDA,丙二醛;MFC-Gem,载吉西他滨的碳质纳米粒子;MGMT,甲基鸟嘌呤甲基转移酶;MMNPs,磁性介孔二氧化硅纳米粒子;MMP-2,金属蛋白酶-2;MnFe 2 O 4 ,锰铁氧体;mRNAs,信使 RNA;NEPC,神经内分泌前列腺癌;NF- κ B,活化 B 细胞的核因子 κ 轻链增强子;NFS1,半胱氨酸脱硫酶;NK,自然杀伤细胞; NOX,NADPH 氧化酶 1;NRF2,核因子红细胞 2 相关因子 2;NSCLC,非小细胞肺癌;OC1,耳蜗毛细胞;OS,总生存率;P62,隔离小体 1;PET,正电子发射断层扫描;P-GP,P-糖蛋白;PCC,持久癌细胞;PCN(Fe) MOFs,Fe 3 + 卟啉金属有机骨架上的 PEG;PD-L1,程序性死亡配体 1;PDAC,胰腺导管腺癌;PEG,聚乙二醇;PGE2,前列腺素 E2;PGRMC1,孕酮受体膜成分 1;PHPM,ROS 敏感聚合物;PTX,紫杉醇;PUFA,多不饱和脂肪酸;PUFA-OOH,磷脂多不饱和脂肪酸过氧化物;RIPK-1/2/3,受体相互作用丝氨酸/苏氨酸蛋白激酶 1/2/3;ROS,活性氧;RR,反应率;siRNA,小干扰 RNA;siSLC7A11,SLC7A11 siRNA;SLC3A2,溶质载体家族 3 成员 2;SLC40A1,溶质载体家族 40 成员 1;SLC7A11,溶质载体家族 7 成员 11;STAT1,信号转导和转录激活因子 1;TAM,肿瘤相关巨噬细胞;TCA,三羧酸循环;TFR,转铁蛋白受体;TME,肿瘤微环境; TMZ,替莫唑胺;TP53,细胞肿瘤抗原 p53;TRADD,肿瘤坏死因子受体 1 型相关死亡结构域蛋白;TTP,进展时间;US FDA,美国食品药品管理局;UTRs,非翻译区;VDAC,电压依赖性阴离子通道;xCT,谷氨酸-胱氨酸反向转运蛋白;Z-VAD-FMK,羧苄氧缬氨酰丙氨酰天冬氨酰-[O-甲基]-氟甲基酮;γ-GCS,γ-谷氨酰半胱氨酸合成酶。 * 通讯作者。电子邮箱地址:mateusz.kciuk@biol.uni.lodz.pl (M. Kciuk)。