XiaoMi-AI文件搜索系统
World File Search System过长
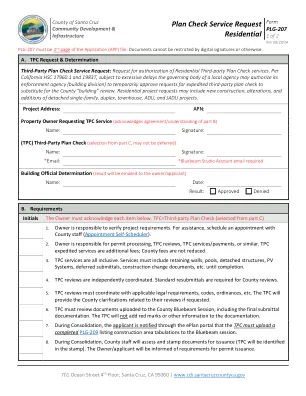
计划检查服务请求住宅
第三方计划检查服务请求:请求授权住宅第三方计划检查服务。根据加州 HSC 17960.1 和 19837,如果延误时间过长,当地机构的管理机构可以授权其执法机构(建筑部门)暂时批准加快第三方计划检查的请求,以代替县“建筑”审查。住宅项目请求可能包括新建、改建和增建独立单户住宅、复式住宅、联排别墅、ADU 和 JADU 项目。


小狗的社会化和疫苗接种
社会化是将幼犬引入世界的过程,帮助它建立自信,不惧怕一生中遇到的人、地方和事物。最关键的社会化窗口发生在 12 周龄之前,该窗口被认为在 5 个月时关闭。通常,幼犬进行社会化时的年龄越小,它获得自信的速度就越快、越容易。等待时间过长可能会导致需要更困难的康复过程,而不是社会化。大多数攻击性问题只是社会化效率低下的结果。
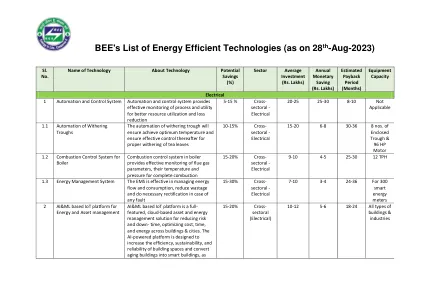
Bee的节能技术清单(如28-... 印度能源方案 开发自愿设计的蓝图... 能源效率措施的影响
在火电管锅炉(两通道和三通)中,热燃烧气体通过长而小型的管,在那里热量通过管墙传递到水中。Firetube锅炉按其“通行证”数量或热燃烧气体穿过锅炉热交换表面的次数进行分类。湍流器可以是降低堆栈温度并提高单通气水平回流管(HRT)砖锅锅炉以及较早的两种和三通油和天然气和天然气燃料的火器锅炉的燃料到蒸汽效率的一种成本效益方法。

医院战略 2019 – 2024 | 人们关爱……
TUH 最初设计为拥有 650 张床位的医院,但由于当时预算限制,最终建成时减少了 120 张床位。自开业以来,医院的服务需求不断增长,但容量却没有相应增加。这导致患者等待时间过长。医院迫切需要提高容量和新的工作方式,以满足日益增加和更复杂的需求。幸运的是,与许多其他医院不同,TUH 并不局限于其 33 英亩的校园,能够横向和纵向扩展以及异地扩展,成为一家“无墙医院”。

2022 年春/夏 - 国防旅行管理办公室
在直接联系航空公司之前,您必须尝试联系 TMC。您只能在 TMC 无法提供服务或等待时间过长时联系航空公司。如果您原来的航空公司无法“保护/重新预订”您的机票,您可以直接向其他航空公司预订。(请务必保留您原始未使用的机票号码的记录。)申请预订美国政府机票(如果有),并确保您可以访问您的旅行订单(如果适用)和有效的旅行卡。必须使用最低成本的经济舱或经济舱票价住宿。任何升级的住宿都必须按照 JTR 第 020206-J 段的规定获得授权或批准。

共同过渡到远程学习 - 生命大学
时间分块法 远程学习时保持高效并减少干扰的方法之一是时间分块法。考虑将任务分成 20 分钟到 1 小时不等的时间段。在这段时间内,专注于一个主题领域或作业,并减少所有其他干扰。这意味着在讲课期间关闭所有其他标签、关闭手机或将手机调至“飞行模式”、不要查看电子邮件和/或在门上挂上“请勿打扰”的标志,以免家人或室友打扰。这段时间结束后,你可以休息 5-10 分钟来奖励自己。但是,休息时间也不宜过长,因为这样可能很难激励自己重新开始工作。

引力波 - ijrpr
探测引力波的挑战在于它们在时空中造成的极小扭曲,而这些扭曲很容易被环境噪声掩盖。克服这些挑战需要先进的技术来降低地震活动、热波动和其他来源的噪声(Abbott 等人,2016 年)。一些关键策略包括:首先,地震隔离:LIGO、Virgo 和 KAGRA 中的悬挂镜被设计为与地面振动隔离。多层悬挂系统(包括主动阻尼机制)有助于保护镜子免受地震干扰(Thorne,2017 年);其次,真空系统:这些探测器中的激光束穿过长真空管,以防止空气分子散射,从而将噪声引入测量中。

人工智能与数据隐私:平衡紧张关系
虽然 ICO 制定的指南包含大量有用信息,但往往篇幅过长,有时内容重叠,且起草风格不一致。因此,组织可能很难知道从哪里开始,或者各个指南如何组合在一起。但是,去年 11 月,它发布了有关如何更好地使用 AI 的提示(如下所述)。本指南相对较短且内容丰富,使其成为组织更易于访问的起点。ICO 的 AI 风险缓解工具包于去年完成,也旨在为组织提供更实用的支持。这两种新资源还链接到更详细指南的相关部分。除了指南和工具包之外,ICO 的监管沙盒也是 AI 公司的有用资源。

人工智能与神经网络简介
数据集不具代表性会导致模型在训练数据集上表现良好,但在实际使用中,当输入新数据或独特数据时,模型无法泛化。然而,原因却不同。如果模型训练时间过长或具有过多容量,用于记忆训练数据(而不是识别其中的潜在模式)或学习其中不相关的模式,则会发生过度拟合。例如,电子邮件垃圾邮件检测模型可能会在训练期间记住标记为垃圾邮件的电子邮件中的特定单词或短语。当这些确切的单词没有出现在现实世界的垃圾邮件中时,它就无法将其检测为垃圾邮件。