XiaoMi-AI文件搜索系统
World File Search System逆行
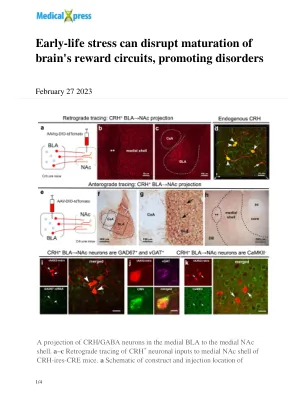
早期的压力会破坏大脑奖励电路的成熟,促进疾病
AAV2 -RETRO -CAG -FLEX -TDOMATO -WPRE病毒,可允许逆行进入投影神经元,从而为NAC提供传入的输入。b示例局部感染的CRH +轴突末端的共聚焦显微照片在内侧NAC壳中。c逆行跟踪将内侧BLA识别为CRH + NAC输入的强大来源。d 3D图像(z-stack;0.5μm步长)确认在AAV-RETRO感染细胞(红色)的BLA中定位,共表达内源性CRH(绿色);双重标记的神经元=黄色。e - 从BLA到内侧NAC壳的CRH +轴突投影的g顺行追踪。e,AAV1-DIO-TDTOMATO构建体和病毒遗传实验设计。f病毒注射仅限于中央杏仁核(CEA)的BLA,G,通过在BLA CRH +神经元中的TDTomato选择性表达显示。h bla-origin CRH +轴突和内侧NAC壳中的端子。i - K病毒注射到BLA中的内侧NAC壳逆转录感染的SOMATA。i将荧光原位杂交(FISH)与BLA中CRH +细胞中的GAD67 mRNA进行免疫染色。箭头指向共定位的GAD67 mRNA和病毒 - 重复蛋白标记。j a bla→NAC细胞(红色)共表达内源性CRH(绿色)和VGAT(Magenta),但K不会共表达谷氨酸能标记Camkii。** = Calleja的主要岛,AC前委员会,DB对角线带。i和k中的比例尺= 10 µm。在至少两个独立的垃圾中评估了在小鼠中评估发现发现,病毒注射,投射评估和免疫组织化学的。信用:自然通讯(2023)。doi:10.1038/s41467-023-36780-x

鸭肠炎病毒 UL21 是一种晚期基因,编码一种与 pUL16 相互作用的蛋白质
背景:pUL21 是 Alphaherpesvirinae 的保守蛋白,具有多种重要功能。该亚家族其他成员的 pUL21 的 C 端具有 RNA 结合能力;该结构域有助于伪狂犬病毒 (PRV) 体外和体内逆行轴突运输,并参与新复制的病毒 DNA 包装和细胞内病毒运输。然而,关于鸭肠炎病毒 (DEV) pUL21 的知识有限。结果:我们证实 DEV UL21 是一个编码结构蛋白的 γ 2 基因。此外,我们观察到 pUL21 定位于细胞核和细胞质中。DEV pUL21 与 pUL16 相互作用并在转染的人胚胎肾 (HEK) 293 T 细胞和 DEV 感染的鸭胚胎成纤维细胞 (DEF) 中形成复合物。这些结果通过 CO-IP 测定得到进一步证实。

使用磁力计和光传感器进行定位
第一种定位技术基于一个或多个磁力计测量磁性物体的感应磁场。这些测量取决于物体的位置和磁特征,可以用从电磁理论中得出的模型来描述。对于这项技术,已经分析了两种应用。第一个应用是交通监控,它对强大的定位系统有很高的需求。通过在车道附近部署一个或多个磁力计,可以检测和分类车辆。这些系统可用于安全目的,例如检测高速公路上的逆行驾驶员,以及通过监测交通流量用于统计目的。第二个应用是室内定位,其中移动磁力仪测量室内环境中磁结构引起的静止磁场。在这项工作中,提出并评估了此类磁环境的模型。

一例引人注目的记忆障碍病例的神经病理学为人类记忆提供了启示
肯特·科克伦 (KC) 在 30 岁时因摩托车事故导致颅内创伤,导致其出现严重的失忆症,此后近三十年来研究人员一直在对他进行研究。KC 在言语和非言语领域均患有严重的前行性遗忘症,同时伴有选择性逆行性遗忘症,无法记住受伤前经历的个人事件(情景记忆),而对个人和世界事实的记忆(语义记忆)以及内隐记忆则相对保留。这种保留和受损的记忆模式延伸到对大规模环境的空间记忆,以及对未来的想象和决策。62 岁时死后大脑检查结果包括中度弥漫性萎缩、左侧眶额挫伤、左侧大脑后动脉梗塞和左前额分水岭梗塞。值得注意的是,双侧海马均有严重的神经元丢失和神经胶质增生。左侧海马前部和后部受到严重影响,但 CA2、CA4 和齿状回 (DG) 局部未受损。左侧穹窿伴有退化。右侧海马前部几乎完全破坏,后部相对保存,主要是 CA4 和 DG。双侧海马旁回和左侧丘脑前部也出现神经元丢失和神经胶质增生。β-淀粉样蛋白、磷酸化 tau 或 TDP-43 免疫染色未发现共存的神经退行性现象。内侧颞叶结构受损程度与 KC 严重的顺行性和逆行性遗忘症相符,但意外发现 CA2/CA4 和 DG 保存完好。 KC 的案例表明,在广泛的脑损伤后,相对明显的功能分离仍然可能存在,结构受损的脑区不太可能对完整的认知功能至关重要。这样一来,本文提出的研究结果为 KC 对我们理解记忆的临床解剖关系做出了重要贡献。

吸力是内在学的未来吗? eau ...
从光纤到数字系统的过渡显着增强了术中视图。1此外,引入一次性范围和范围持续的小型化还具有进一步的内窥镜石材手术。现代激光技术的利用,尤其是:Yag Laser(HO:YAG)和Thulium纤维激光片质脆性,显着为改善的手术结果做出了贡献。逆行肾脏内手术(RIRS)被当前指南推荐,作为包括大于2 cm的石头在内的肾结石的第一或第二选择。2尽管采用了这些技术进步的优势,但由于雪球效应或残留片段(RFS)引起的互操作观点(RFS),取得较大的微积分的成功率可能是具有挑战性的。我们的目标是完全无石的速度状态,但RFS的主张仍然是一个关注的问题,通常需要进行额外的干预措施和更多的卫生保健系统成本。3尽管已经提出了临床上微不足道的RF的概念,但没有一个
奇塔兰詹国家癌症研究所
RD-12 X 射线脊柱 AP 0 50 400 RD-13 X 射线脊柱侧位 0 50 400 RD-14 X 射线骨盆 AP 0 50 400 RD-15 X 射线骨盆侧位 0 50 400 RD-16 X 射线颈部 AP 0 50 400 RD-17 X 射线颈部侧位 0 50 400 RD-18 X 射线上肢/关节 AP 0 50 400 RD-19 X 射线上肢/关节侧位 0 50 400 RD-20 X 射线下肢/关节 AP 0 50 400 RD-21 X 射线下肢/关节侧位 0 50 400 RD-22 X 射线胸部 PA 0 50 400 RD-23 X 光胸部侧位 0 50 400 RD-24 X 光腹部 0 50 400 RD-25 X 光腹部平片 0 50 400 RD-26 X 光骨骼检查 0 800 4400 RD-27 X 光便携式 0 100 650 RD-28 X 光PNS 0 50 480 RD-29 X 光胸骨AP 0 50 480 RD-30 X 光胸骨斜位 0 50 480 RD-31 X 光胸骨侧位 0 50 480 对比 NC GP RD-32 X 光涎管造影 0 250 1400 RD-33 X 光钡餐检查 0 250 1400 RD-34 X 射线 康雷吞咽 350 700 1400 RD-35 X 射线 钡餐 0 350 1900 RD-36 X 射线 钡餐跟进 0 650 4000 RD-37 X 射线 钡灌肠 0 650 4000 RD-38 X 射线 小碗灌肠 0 650 4000 RD-39 X 射线 胆管造影 0 150 800 RD-40 X 射线 ERCP 0 850 4960 RD-41 X 射线 IVP 0 500 2800 RD-42 X 射线 膀胱造影 0 250 1400 RD-43 X 射线 MCU 0 300 2000 RD-44 X 射线逆行尿道造影 0 250 1400 RD-45 X 射线逆行肾盂造影 0 250 1400 RD-46 X 射线正中造影 0 150 1000

使用星形胶质细胞神经网络进行容错计算的设计方法
我们提出了一种设计方法来促进深度学习模型的容错。首先,我们实现了一个多核容错神经形态硬件设计,其中每个神经形态核心中的神经元和突触电路都包裹在星形胶质细胞电路中,星形胶质细胞是大脑的星形神经胶质细胞,它通过使用闭环逆行反馈信号恢复故障神经元的尖峰放电频率来促进自我修复。接下来,我们在深度学习模型中引入星形胶质细胞,以实现对硬件故障所需的容忍度。最后,我们使用系统软件将支持星形胶质细胞的模型划分为集群,并在所提出的容错神经形态设计上实现它们。我们使用七种深度学习推理模型评估了这种设计方法,并表明它既节省面积又节能。

双重麻烦:在心力衰竭的情况下发生不断发展的腿部溃疡的情况
静脉腿溃疡(VLU)是晚期慢性静脉功能不全(CVI)的表现。在高级阶段,慢性静脉功能不全导致静脉瓣膜无能,这会导致逆行血流,阻塞或两者兼而有之。良好的静脉功能不全与心脏异常有关。尽管慢性心力衰竭(CHF)是一种普遍的心脏病,被认为是发展静脉溃疡的危险因素,但最近的研究发现两种疾病之间没有双向因果关系,并且建议进一步研究[1]。本报告提出了先前一致改善静脉溃疡,在急性CHF加剧过程中表现出停滞和恶化。溃疡被感染,并且愈合过程受到阻碍,从而促使手术清创术。此病例表明,急性CHF加剧可能在静脉溃疡代偿上发挥作用,并使两种病理学之间因果关系的可能性提供了信誉[2]。

空降战术的发展(1935-2020)
到了 20 世纪 30 年代,飞机和降落伞的广泛使用使得通过降落伞将轻型步兵部署到敌后成为可能。陆军野战手册 FM 3-90(空降和空中突击作战)的附录 C 涉及空降作战和伞兵。4 伞兵作战在“空降作战”一节中讨论。空降作战的战术梯队包括:突击梯队、后续梯队和后卫梯队。在占领确定空中前沿的目标后,突击梯队确保空降部队、装备和补给的畅通无阻。后续梯队可以包括轻型和重型联合部队。根据指挥官的决定,重型武器可以通过降落伞或空降方式投送。由于防空技术的进步,到 20 世纪 50 年代末,大规模部署空中突击部队的风险越来越大。同时,燃气涡轮运输机运输能力的提高使得空运轻型坦克和自行火炮等重型装备成为可能。随着先进降落伞货物运输系统(低空降落伞提取系统、逆行火箭等)的发展,空降部队增加了一些重型组织单位(例如装甲营),从而提高了战斗力和机动性。从 1980 年代开始,美国第 82 空降师(

Rothamsted存储库下载
抽象的植物细胞经常遇到正常生长和发育的一部分,或响应诸如洪水等环境压力的一部分。近年来,我们对低氧反应基因表达的多层控制的理解已大大增加。在此更新中,我们对调节对低氧水平的反应的表观遗传,转录,翻译和翻译后机制进行了广泛的看法。我们强调了翻译后修饰(包括磷酸化),次级信使,转录级联反应以及来自线粒体和网状网状(ER)的逆行信号如何如何控制转录因子活性和低氧基因转录的控制。我们讨论了通过专注于主动和抑制性的染色质修饰和DNA甲基化的表观遗传机制,以调节对氧气供应减少的反应。我们还描述了当前对紧密调节mRNA翻译以协调缺氧下有效基因表达的共同和转录机制的知识。最后,我们在该领域提出了一系列杰出的问题,并考虑了如何对低氧触发的监管层次结构的分子起作用的新见解,这可能为开发洪水的作物铺平道路。