XiaoMi-AI文件搜索系统
World File Search System驱动电流

CoolSiC™ MOSFET 栅极驱动电压窗口指南
3.1 1200 V 器件准则 ...................................................................................................................................... 7
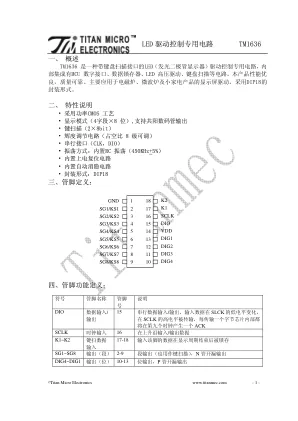
LED 驱动控制专用电路TM1636
在有按键按下时,读键数据如下: SG1 SG2 SG3 SG4 SG5 SG6 SG7 SG8 K1 1110_1111 0110_1111 1010_1111 0010_1111 1100_1111 0100_1111 1000_1111 0000_1111 K2 1111_0111 0111_0111 1011_0111 0011_0111 1101_0111 0101_0111 1001_0111 0001_0111 在无按键按下时,读键数据为: 1111_1111 ; 七、 接口说明 微处理器的数据通过两线总线接口和 TM1636 通信,在输入数据时当 SCLK 是高电 平时, DIO 上的信号必须保持不变;只有 SCLK 上的时钟信号为低电平时, DIO 上的信号 才能改变。数据输入的开始条件是 SCLK 为高电平时, DIO 由高变低;结束条件是 SCLK 为高时, DIO 由低电平变为高电平。 TM1636 的数据传输带有应答信号 ACK ,在传输数据的过程中,在时钟线的第九个 时钟芯片内部会产生一个应答信号 ACK 将 DIO 管脚拉低。 指令数据传输过程如下图(读按键数据时序):

LED 驱动控制专用电路TM1637
符号 单位: 毫米 最小值 典型值 最大值 A 3.71 4.00 4.31 A1 0.50 0.60 0.80 A2 3.20 3.40 3.60 B 0.33 0.45 0.53 B1 1.525(TYP) C 0.20 0.28 0.36 D 25.70 26.00 26.54 E 6.20 6.40 6.75 E1 7.32 7.78 8.25 e 2.54(TYP) L 3.00 3.30 3.60 E2 8.20 8.70 9.10 B2 0.87 1.02 1.17

LED 显示屏行驱动芯片TM3301
集成串行译码电路 集成 8 高效 PMOS 输出 , 导通电阻 100mΩ 集成内部防烧功率管 动态消影技术 反向击穿保护 支持最大持续电流 2.5A 低功耗设计 消影电位 8 档可调 封装形式: SOP16 广泛应用领域: LED 显示屏、 LED 照明、 LED 景观亮化

LCD 驱动控制专用电路TM1723
Bit0 Bit1 Bit2 Bit3 Bit4 Bit5 Bit6 Bit7 位 图( 3 ) ▲注意: 1 、 TM1723 最多可以读 2 个字节,不允许多读。 2 、读数据字节只能按顺序从 BYTE1-BYTE2 读取,不可跨字节读。例如:硬件上的 KEY2 与 KS3 对应按键按下时, 此时想要读到此按键数据,必须需要读到第 2 个字节的第 6BIT 位,才可读出数据;当 KEY1 与 KS3 , KEY2 与 KS3 , KEY3 与 KS3 三 个按键同时按下时,此时 BYTE2 所读数据的 B5 , B6 , B7 位均为 1 。 3 、组合键只能是同一个 KS ,不同的 KEY 引脚才能做组合键;同一个 KEY 与不同的 KS 引脚不可以做成组合键使用。 7.3.按键扫描

32*8 & 24*16 LED 驱动芯片TM1681
TM1681 的系统时钟用来产生系统工作的时钟频率。LED 驱动时钟、系统时钟可以取自片内的 RC 振 荡器(256KHz)或者使用 S/W 设置由外部时钟输入。系统振荡器构造如图7 所示。当SYS DIS 命令被 执行时,系统时钟停止,LED 工作循环将被关闭(这条指令只能适用与片内 RC 振荡器)。一旦系统时 钟停止时,LED 显示为空白,时基也会丧失其功能。LED_OFF 命令用来关闭 LED 工作循环,LED 工作 循环被关闭之后,用 SYS DIS 命令节省电源开支,充当省电命令;如果是片外时钟源被选择的话,使 用 SYS DIS 命令不能够关闭振荡器以及执行省电模式。晶体振荡器可以通过OSC 管脚提供时钟频率, 在这种情况下,系统将不能进入省电模式。在系统上电时,TM1681 默认处在 SYS DIS 状态下。
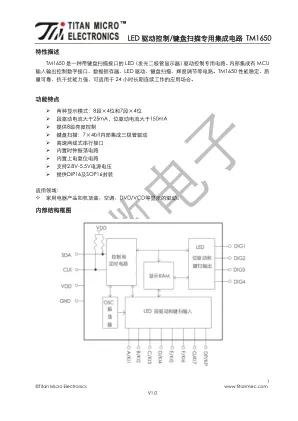
LED 驱动控制/键盘扫描专用集成电路TM1650
接口和TM1650 通信,在输入数据时当SCL 是高电平时,SDA 上的信号必须保持不变;只有SCL 上的 时钟信号为低电平时,SDA 上的信号才能改变。数据输入的开始条件是SCL 为高电平时,SDA 由高变
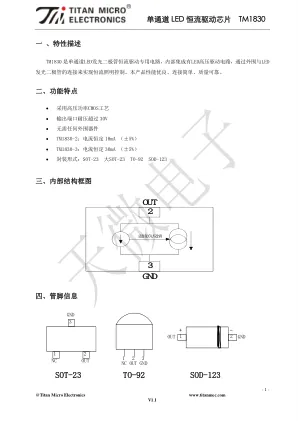
单通道LED 恒流驱动芯片TM1830
如图1 所示,要使TM1830 工作在恒流状态下,芯片OUT 引脚上电压应大于2.2V,即芯片的2、3 脚之间的电压应达到2.2V 以上。在应用时,电源串接LED 灯后加在OUT 引脚上的电压建议在3.0V 左右。 如果芯片持续工作在额定恒流状态下,TM1830-2 和TM1830-3 的OUT 引脚电压应分别在12.0V 和8.0V 以内为宜。

微生物硫代谢及其驱动下建立的生物生态关系
微生物,动物和植物中的代谢途径表现出各种关系。基于微生物硫代谢,本文总结了微生物,动物和植物中硫的四个主要代谢途径,并强调了相似性,差异和关系。微生物是生物硫循环的主要驱动力,参与硫的所有主要代谢途径。微生物通过微生物减少了硫磺硫,可减少甲烷在环境中的挥发。微生物或植物的同化硫还原性的动物有机硫来源,而动植物则缺乏异化或同化硫还原的功能。硫氧化发生在所有三种生物体中,具有相似的途径,其中硫转移酶多样化氧化产物。植物中的硫矿化尚不清楚,但是动物或微生物的矿化使植物中的硫硫底物可促进其他无机硫底物。 在本质上,基于硫代谢的生态关系,例如肠道微生物与宿主动物之间的关系,根际微生物与植物根,衰减的动物和植物的微生物矿化,以及微生物氧化的微生物矿化,硫磺的硫化和减少,显着增强了硫磺的硫磺含量。硫矿化尚不清楚,但是动物或微生物的矿化使植物中的硫硫底物可促进其他无机硫底物。在本质上,基于硫代谢的生态关系,例如肠道微生物与宿主动物之间的关系,根际微生物与植物根,衰减的动物和植物的微生物矿化,以及微生物氧化的微生物矿化,硫磺的硫化和减少,显着增强了硫磺的硫磺含量。

分子运动模型中的空间驱动电流逆转
模拟可以帮助揭示分子结构决定功能的复杂方式。在这里,我们使用分子模拟来说明分子电动机结构的轻微变化会导致电动机的典型动力学行为与反向方向相反。受自主合成链苯烷电动机的启发,我们研究了最小运动模型的分子动力学,由沿着沿着包含散布的结合位点和催化位点的轨道移动的穿梭环组成。结合位点吸引了穿梭环,而催化位点加快了分子物种之间的反应,可以将其视为燃料和废物。当将燃料和废物保持在非平衡稳态浓度中时,反应驱动沿轨道的飞行环的指示运动的自由能。使用此模型和非平衡分子动力学,我们表明可以通过简单地调整轨道上的结合和催化位点之间的间距来逆转穿梭环的方向。我们提出了当前逆转背后的一个空间机制,并由模拟的动力学测量支持。这些结果证明了分子模拟如何指导人工分子电机的未来发展。