XiaoMi-AI文件搜索系统
World File Search System齐萨基

健康的基本生理和生化研究
ACTN3 R577X多态性。 J锻炼营养生物化学。 2015; 19(3):157-64。 3 Kikuchi N,Yoshida S,Min SK,Lee K,Sakamaki-Sunaga M,Okamoto T等。 ACTN3 R577X基因型与日本人群中的肌肉功能有关。 Appl Physiol Nutr Metab。 2015; 40(4):316-22。 4 Gatfield D,Izaurralde E.胡说八道介导的信使RNA衰变是由果蝇的核核酸裂解引发的。 自然。 2004; 429(6991):575-8。 5 Tuladhar R,Yeu Y,Tyler Piazza J,Tan Z,Rene Clemenceau J,Wu X等。 基于CRISPR-CAS9的诱变经常引起目标mRNA的正调。 nat Commun。 2019; 10(1):4056。ACTN3 R577X多态性。J锻炼营养生物化学。2015; 19(3):157-64。3 Kikuchi N,Yoshida S,Min SK,Lee K,Sakamaki-Sunaga M,Okamoto T等。 ACTN3 R577X基因型与日本人群中的肌肉功能有关。 Appl Physiol Nutr Metab。 2015; 40(4):316-22。 4 Gatfield D,Izaurralde E.胡说八道介导的信使RNA衰变是由果蝇的核核酸裂解引发的。 自然。 2004; 429(6991):575-8。 5 Tuladhar R,Yeu Y,Tyler Piazza J,Tan Z,Rene Clemenceau J,Wu X等。 基于CRISPR-CAS9的诱变经常引起目标mRNA的正调。 nat Commun。 2019; 10(1):4056。3 Kikuchi N,Yoshida S,Min SK,Lee K,Sakamaki-Sunaga M,Okamoto T等。ACTN3 R577X基因型与日本人群中的肌肉功能有关。Appl Physiol Nutr Metab。2015; 40(4):316-22。4 Gatfield D,Izaurralde E.胡说八道介导的信使RNA衰变是由果蝇的核核酸裂解引发的。自然。2004; 429(6991):575-8。5 Tuladhar R,Yeu Y,Tyler Piazza J,Tan Z,Rene Clemenceau J,Wu X等。基于CRISPR-CAS9的诱变经常引起目标mRNA的正调。nat Commun。2019; 10(1):4056。

萨蒂什·萨贾
Google Scholar https://scholar.google.com/citations?user=DjYlusAAAAAJ&hl=en Scopus https://www.scopus.com/authid/detail.uri?authorId=57195424605 ORCID http://orcid.org/0000-0002-6604-4095 Publons https://publons.com/researcher/2227602/satish-sajja/ LinkedIn https://www.linkedin.com/in/satish-sajja-29616b18/ 咨询/行业项目



CRISPR-CasとOMEGashisutemuの分子基盘 - 生化学
202. 3) Wang, JY, Tuck, OT, Skopintsev, P., Soczek, KM, Li, G., Al-Shayeb, B., Zhou, J., & Doudna, JA (2023) 通过 CRISPR 修剪器整合酶进行基因组扩展。Nature,618,855 ‒ 861。4) Wang, JY, Pausch, P., & Doudna, JA (2022) CRISPR-Cas 免疫和基因组编辑酶的结构生物学。Nat. Rev. Microbiol. , 20 , 641 ‒ 656。5) Anzalone, AV、Randolph, PB、Davis, JR、Sousa, AA、Ko-blan, LW、Levy, JM、Chen, PJ、Wilson, C.、Newby, GA、Raguram, A. 等人 (2019) 无需双链断裂或供体 DNA 的搜索和替换基因组编辑。Nature,576,149 ‒ 157。6) Mehta, J. (2021) CRISPR-Cas9 基因编辑用于治疗镰状细胞病和β地中海贫血。N. Engl. J. Med.,384,e91。 7) Kapitonov, VV, Makarova, KS, & Koonin, EV (2015) ISC,一组编码 Cas9 同源物的新型细菌和古细菌 DNA 转座子。J. Bacteriol. ,198,797 ‒ 807。8) Altae-Tran, H., Kannan, S., Demircioglu, FE, Oshiro, R., Nety, SP, McKay, LJ, Dlakić, M., Inskeep, WP, Makarova, KS, Macrae, RK, et al. (2021) 广泛分布的 IS200/IS605 转座子家族编码多种可编程的 RNA 引导的核酸内切酶。 Science , 374 , 57 œ 65。9) Weinberg, Z., Perreault, J., Meyer, MM, & Breaker, RR (2009) 细菌宏基因组分析揭示的特殊结构化非编码 RNA。Nature , 462 , 656 œ 659。10) Hirano, S., Kappel, K., Altae-Tran, H., Faure, G., Wilkinson, ME, Kannan, S., Demircioglu, FE, Yan, R., Shiozaki, M., Yu, Z., et al. (2022) OMEGA 切口酶 IsrB 与 ω RNA 和靶 DNA 复合的结构。 Nature , 610 , 575 œ 581。11) Biou, V., Shu, F., 和 Ramakrishnan, V. (1995) X 射线晶体学显示翻译起始因子 IF3 由两个通过 α 螺旋连接的紧凑的 α/β 结构域组成。EMBO J. , 14 , 4056 œ 4064。12) Schuler, G., Hu, C., 和 Ke, A. (2022) IscB-ω RNA 进行 RNA 引导的 DNA 切割的结构基础以及与 Cas9 的机制比较。 Science,376,1476 ‒ 1481。13) Bravo, JPK、Liu, MS、Hibshman, GN、Dangerfield, TL、Jung, K.、McCool, RS、Johnson, KA 和 Taylor, DW (2022) CRISPR-Cas9 错配监测的结构基础。Nature,603,343 ‒ 347。14) Aliaga Goltsman, DS、Alexander, LM、Lin, JL、Fregoso Ocampo, R.、Freeman, B.、Lamothe, RC、Perez Rivas, A.、Temoche-Diaz, MM、Chadha, S.、Nordenfelt, N. 等人 (2022) 从未培养的微生物中发现用于基因组编辑的紧凑型 Cas9d 和 HEARO 酶。Nat. Commun. ,13,7602。
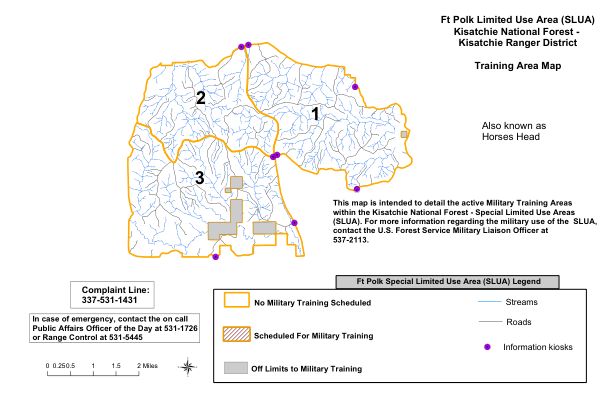
波尔克堡有限使用区 (SLUA) 基萨奇国家森林
该地图旨在详细说明基萨奇国家森林 - 特殊限制使用区域 (SLUA) 内的现役军事训练区。有关 SLUA 军事用途的更多信息,请联系美国森林服务局军事联络官,电话:537-2113。

痴呆症的语言标记及其在阿尔茨海默氏病早期诊断中的作用:通过句子探索语法和句法能力
塞洛尼基亚里士多德大学,塞萨洛尼基亚里士多德大学英语学院,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基大学医学院,塞萨洛尼基大学,塞萨洛尼基大学,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基,希腊阿尔茨阿尔茨阿尔兹·阿尔茨·阿尔兹·阿尔兹·艾尔兹·迪克(Greece)的泰士(Grecect)塞萨洛尼基的技术赫拉斯,希腊语言学系,语言学系,亚里士多德大学塞萨洛尼基大学哲学学院塞萨洛尼基(Thessaloniki),塞萨洛尼基(Thessaloniki),希腊h神经退行性疾病实验室,跨学科研究与创新中心(CIRI - AUTH),巴尔干中心,塞萨洛尼基亚里士多德大学,塞萨洛尼基,塞萨洛尼基,格里西基,格里西基,塞洛尼基亚里士多德大学,塞萨洛尼基亚里士多德大学英语学院,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基大学医学院,塞萨洛尼基大学,塞萨洛尼基大学,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基,塞萨洛尼基,希腊阿尔茨阿尔茨阿尔兹·阿尔茨·阿尔兹·阿尔兹·艾尔兹·迪克(Greece)的泰士(Grecect)塞萨洛尼基的技术赫拉斯,希腊语言学系,语言学系,亚里士多德大学塞萨洛尼基大学哲学学院塞萨洛尼基(Thessaloniki),塞萨洛尼基(Thessaloniki),希腊h神经退行性疾病实验室,跨学科研究与创新中心(CIRI - AUTH),巴尔干中心,塞萨洛尼基亚里士多德大学,塞萨洛尼基,塞萨洛尼基,格里西基,格里西基,


