XiaoMi-AI文件搜索系统
World File Search System串扰

毛果杨体外转化的全基因组关联研究和网络分析支持多种植物激素途径和串扰的关键作用
许多植物物种和基因型对转化和再生 (TR) 的适应性存在很大差异,这对基因工程在研究和育种中的应用提出了挑战。为了帮助了解这种变异的原因,我们使用 1204 棵野生黑杨树种群进行了关联作图和网络分析。为了对愈伤组织和嫩枝 TR 进行精确和高通量的表型分析,我们开发了一种计算机视觉系统,可以交叉引用互补的红、绿、蓝 (RGB) 和荧光高光谱图像。我们使用单标记和组合变异方法进行了关联作图,然后对已发表的多组学数据集进行了上位性和整合的统计检验,以确定可能的调控中心。我们报告了 409 个与编码序列 5 kb 范围内的关联有关的候选基因,上位性测试表明其中 81 个候选基因是彼此的调节因子。与蛋白质 - 蛋白质相互作用和转录调控相关的基因本体术语被过度使用。除了长期确定对 TR 至关重要的生长素和细胞分裂素通路之外,我们的结果还强调了应激和伤害通路的重要性。这些通路内和跨通路的潜在信号调节中心包括生长调节因子 1 (GRF1)、磷脂酰肌醇 4-激酶 β 1 (PI-4K β 1) 和 OBF 结合蛋白 1 (OBP1)。

癌症干细胞和与肿瘤相关的巨噬细胞作为肿瘤进展的伴侣:串扰和晚期生物信息学工具的机制,以剖析其表型和相互作用
癌症干细胞(CSC)是肿瘤质量中的一个小子集,这显着促进了癌症的进展,通过各种致癌途径的失调,促进肿瘤生长,化学抗性和转移形成。CSC的侵略性行为由几种细胞内信号通路,例如Wnt,NF-KAPPA-B,Notch,HydgeHog,Jak-Stat,Pi3K/Akt1/MTOR,TGF/TGF/TGF/SMAD,PPAR,PPAR,PPAR和MAPK激酶,以及诸如外胞外小叶等信号,以及诸如外胞外叶子,以及分类的cy虫,以及分类的分解。趋化因子,促血管生成和生长因子,最终调节CSC表型。在这种情况下,肿瘤微环境(TME)是建立允许性肿瘤生态位的关键参与者,其中CSC与各种免疫细胞进行复杂的通信。“致癌”免疫细胞主要由B和T淋巴细胞,NK细胞和树突状细胞表示。在免疫细胞中,巨噬细胞由于其不同的亚群而表现出更塑性和适应性的表型,其特征在于免疫抑制和炎症表型。Speci fi cally, tumor-associated macrophages (TAMs) create an immunosuppressive milieu through the production of a plethora of paracrine factors (IL-6, IL-12, TNF-alpha, TGF-beta, CCL1, CCL18) promoting the acquisition by CSCs of a stem-like, invasive and metastatic phenotype.tams已经证明了通过直接配体/受体(例如CD90/CD11b,Lsectin/btn3a3,epha4/ephrin)相互作用与CSC进行通信的能力。另一方面,CSC表现出其影响免疫细胞的能力,创造了有利的微环境,以实现癌症的进展。如今,有趣的是,CSC和TME的双向影响会导致表观遗传重编程,从而维持恶性转化。
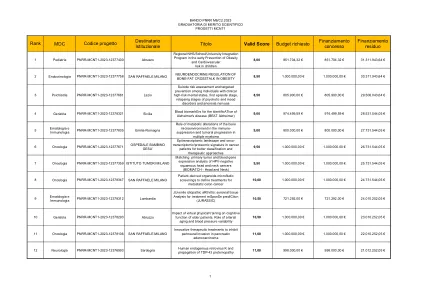
20240315_GRADEPNR2023_MCNT1_SECORNES。
18 Ematologia E Immunologia PNRR-MCNT1-2023-12378434 HUMUNITAS-UP:联合 - 局部串扰性治疗中的链球菌炎12,50 960.000.000.000.000.000.000.000.000.000.000.000,000.000,00,000,000,40.406.406.252,05欧元>>

固态电池:2030 年代的技术,也是 2020 年代的研究挑战
通常,锂离子电池的寿命取决于电池内部的化学反应性。锂离子电池会随着时间、使用和长时间暴露在高温下而退化。对于液体电解质,一种这样的退化机制称为“串扰”。串扰通常涉及过渡金属从正极溶解,并迁移到负极,随后中毒。这种中毒会加速液体电解质的降解,最终导致电池故障。SSE 有很大机会保护电极材料免受这种金属迁移和电极中毒的影响。通过使用 SSE 解决此类问题可以延长锂电池的使用寿命,这对于包括汽车在内的许多应用来说都是非常理想的。

第二个孕期胎儿call体长度的参考图:前瞻性横断面研究
逐渐耗尽。此外,它还逐渐消耗海马中的热休克转录因子1,从而对成年海马神经发生产生负面影响。此外,不仅Piezo2-Piezo2 Crosstalk在本体感受性的初级传入终端和由于丢失的Piezo2引发的Huygens同步而逐渐逐渐破坏了ALS,但Piezo2-Piezo1 crosstalk在Peripery上也破坏了。Syndecans,尤其是神经系统中的Syndecan-3,是维持此压电串扰的关键参与者。syndecan-3的检测到的电荷改变变体可能会促进压电串扰的损害,以及对运动神经元和海马的基于质子的信号的进行性损失。kCNA2的变体还可以促进

全电池 LNMO 与 Si/石墨中锂离子电池电解质的 XPS 研究
摘要:对两种不同类型的电解质(共溶剂和多盐)进行了测试,以用于高压 LiNi 0.5 Mn 1.5 O 4 || Si/石墨全电池,并与含碳酸盐的标准 LiPF 6 电解质(基线)进行了比较。在电池的使用寿命内对阳极和阴极进行原位事后 XPS 分析表明,基线电解质的 SEI 和 CEI 不断增长。在共溶剂电解质中循环的电池表现出相对较厚且长期稳定的 CEI(在 LNMO 上),而确定在 Si/石墨上形成了缓慢增长的 SEI。多盐电解质提供更多富含无机物的 SEI/CEI,同时也形成了本研究中观察到的最薄的 SEI/CEI。在基线电解质电池中发现了串扰,其中在阴极上检测到 Si,在阳极上检测到 Mn。观察发现,多盐电解质和共溶剂电解质均能显著减少这种串扰,其中共溶剂最有效。此外,多盐电解质主要在使用寿命末期检测到铝腐蚀,其中阳极和阴极上均有铝。虽然共溶剂电解质在限制串扰方面提供了更优越的界面性能,但多盐电解质提供了最佳的整体性能,这表明界面厚度比串扰发挥了更好的作用。结合它们的电化学循环性能,结果表明多盐电解质为高压电池提供了更好的电极长期钝化。关键词:LNMO-Si/石墨电池、固体电解质界面、SEI、阴极电解质界面、CEI、表面分析、离子液体电解质

通过精细成型的遮光矩阵减轻 122% NTSC 色域的彩色转换微型 LED 显示器的串扰效应
由于微型 LED 芯片具有广视角特性,制造高色域色彩转换微型发光二极管 (LED) 显示器面临的主要挑战之一是相邻像素之间严重的串扰效应。本研究系统地模拟了导致串扰效应的潜在因素。我们观察到,用遮光矩阵 (LBM) 精确填充每个微型 LED 芯片之间的空间可以成为缓解这种风险的有效解决方案。经过仔细研究,证明了压模辅助成型技术是制造 LBM 的有效方法。然而,实验观察进一步表明,微型 LED 表面残留的黑色 LBM 会严重降低亮度,从而影响显示性能。通过采用等离子蚀刻技术有效提取被捕获的光,成功解决了这个问题。最终,开发了一种顶部发射蓝色微型 LED 背光,该背光采用黑色 LBM 精细成型,并与红色和绿色量子点色彩转换层相结合,实现全彩色显示。我们制造的显示器原型的色域可覆盖国家电视标准委员会的122%。

fbxw7 -notch互动组:一种泛素蛋白酶体系统诱导的串扰调节人体组织中的致癌性转化
图1 E3泛素连接酶和SCF型E3连接酶复合物的结构域结构:A,常见的结构是E3泛素连接酶复合酶配合物,介导许多细胞蛋白的靶向降解。In targeting substrate proteins for degradation, ubiquitin is passed from an E1 ubiquitin-activating enzyme to an E2 ubiquitin-conjugating enzyme to the protein substrate, with the final step (ligating ubiquitin to the substrate) catalyzed by an E3 ubiquitin ligase.b,已知SCF复合物是E3连接酶,而SCF型E3连接酶中的每个复合酶都与一组衔接蛋白相互作用,这些衔接蛋白通过特定的蛋白质 - 蛋白质相互作用域募集不同的结合伴侣,例如WD40 repots,例如重复(LRR)(LRR)(LRR),并在protitate sisstrate for Protiate Degradation degradation。这个数字是由作者(N.K.J.)创建的使用网站https://app.biorender.com [校正于2021年4月27日,在第一次在线出版物之后:图2中的一个错字]

电气工程理学硕士(集成电路专业)
和传输线、PCB 和 IC 材料、阻抗匹配、电源分配网络、并行性和串扰。EMI 和 EMC、反射和终端和 IO 焊盘、片上无源器件、ESD 保护管理、IC 和分立元件封装、热考虑、IC 故障和可靠性、微系统封装和应用。实验室教程:使用集总模型的阻抗匹配、使用微带模型的阻抗匹配、EMC 和 TL 不连续性建模和仿真、键合线建模和仿真、电源分配网络建模、并行传输线建模和分析、串扰和噪声分析、EMI/EMC 干扰分析、片上无源器件和角分析。2. RTL 合成和数字后端:HDL(Verilog)编码基础。可合成的 RTL,

组蛋白/DNA修饰与RNA N
n 6-甲基腺苷(M 6 A)是真核信使RNA(mRNA)中最普遍的内部RNA修饰,在转录和转录后水平调节基因表达。M 6 A与其他良好研究的表观遗传修饰(包括组蛋白修饰和DNA修饰)之间的复杂相互作用已被广泛报道。RNA M 6之间的串扰修饰和组蛋白/DNA修饰在建立基因表达的精确和特定微调的染色质状态和无疑对生理和病理过程都产生深远影响。在这篇综述中,我们讨论了RNA M 6的修改与组蛋白/DNA修饰之间的串扰,强调了它们的复杂通信和基于的机制,以使M 6 A基于M 6的生物学相关性具有全面的看法。