XiaoMi-AI文件搜索系统
World File Search System倒位
充分利用 ChatGPT(人工智能)!
外国导游首次上岗时,需要注意以下几点:沟通的重要性:与游客顺畅的沟通至关重要。如果存在语言障碍,说话要慢,重复单词或短语,以确保游客理解。了解游客的需求:游客期望的体验或服务可能因国家或文化而异。事先收集游客信息,提供符合他们需求的旅游服务非常重要。安全考虑:为确保游客安全,避免前往危险或事故多发区域非常重要。发生灾难时,应采取适当措施安全疏散游客。礼貌服务:需要密切关注游客的舒适度。例如,在炎热的日子里,应鼓励游客保持水分,并提醒他们穿舒适的步行鞋。

数量 1 单位 1 名称
(7)其他 A.须在投标开始前提交《资格审查结果通知书》一份。若您已经提交过,则无需再次提交。 若申请人由代表人或其他代理人代为竞投,则其须于竞投开始前提交《授权委托书》。 邮寄投标应清楚写明公司名称、投标日期和时间、投标主题,并用红色写明“投标书已附上”,并于7月24日星期三下午5点之前邮寄到下述地址。此外,投标人还将提前通过邮件收到投标意向通知。 如果您希望参加投标,您必须于7月18日星期四下午3点之前通过传真或其他方式提交市场价格调查文件。 投标人必须提交“驻军使用标准合同”和“投标和合同指南”(可在东部陆军会计团网站https://www.easternarmy.gov/上获取)。「go jp/gsdf/eae/kaikei/eafin/index html」或泷原警备队会计局办公室。 通过提交您的出价,您将被视为承诺遵守“关于排除有组织犯罪集团的承诺”。投标文件中应当包含下列声明作为接受的表示: “本公司(本人(若为个人)、本公司(若为团体))承诺遵守本承诺书中关于排除有组织犯罪的事项”。若拒绝提交上述“排除有组织犯罪承诺事项”的承诺书,则无法参加投标。若在初次投标时有通过邮寄方式提交的投标人,则重新投标的时间如下。

通过 ddPCR 快速、精确地定量大片段 DNA 切除和倒位
重新混合或改编本材料用于任何目的,无需注明原作者。预印本(未经同行评审认证)在公共领域。它不再受版权限制。任何人都可以合法共享、重复使用,版权所有者已将此版本发布于 2020 年 4 月 14 日。;https://doi.org/10.1101/2020.04.13.039297 doi:bioRxiv 预印本

温度相关选择与大扇贝(Pecten maximus)的染色体倒位有关
基因组分化图景(即基因组中不同种群或物种之间差异的分布)越来越多地被描述,以了解自然选择和重组等微进化力量在导致和维持遗传分化方面所起的作用。这方面研究还表明,染色体结构变异是塑造适应性遗传变异图景的重要因素。由于染色体结构变异的普遍性及其固着性质所必需的强大局部适应压力,双壳类软体动物是探索染色体结构变异与局部适应之间关系的理想分类单元。在这里,我们报告了使用最近的染色体水平基因组组装对东北大西洋自然分布范围内的大扇贝 (Pecten maximus) 进行的种群基因组调查。我们报告了至少三个较大的(12 – 22 Mb)染色体倒位,这些染色体倒位与海面温度有关,其频率与中性种群结构形成对比。这些结果强调了重组抑制染色体倒位在局部适应中可能发挥的巨大作用,并提出了一个假设来解释在相对较小的空间尺度上在王扇贝种群中发现的生殖时间差异的维持。
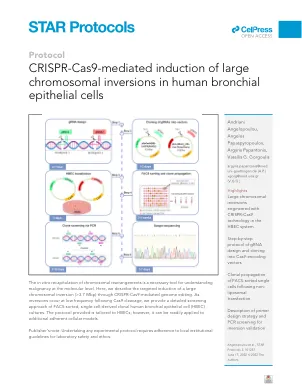
CRISPR-Cas9 介导诱导人类支气管上皮细胞发生大型染色体倒位
1. 通过 UCSC 基因组浏览器 ( https://genome.ucsc.edu/ ) 可获得用于设计两个 gRNA 的目标 DNA 序列。a. 选择感兴趣的基因组版本。在我们的例子中,使用的是“人类 GRCh38/hg38”。b. 根据已知的倒位断点 1 的位置,标记断点前 100-150 bp 到断点后 100–150 bp 范围内的基因组区域。例如,如果断点 1 位于 chr3:2,920,305,则在 UCSC 基因组浏览器搜索框中输入“chr3:2,920,205–2,920,405”以标记所需的染色体区域,然后单击“Go”。c. 在 UCSC 基因组浏览器工具栏上选择“查看”,然后单击“DNA”选项。d.在新窗口中,单击“获取 DNA”以获得准确的 DNA 序列。这是使用 CRISPOR 算法设计 gRNA 引物所需的序列(见下面的步骤 2a)。e. 对倒位的断点 2 重复步骤 1a-1d。2. 要设计 gRNA,请使用 CRISPOR 算法(http://crispor.tefor.net/):a. 输入从步骤 1d 获得的断点 1 的 DNA 序列。确保参考基因组与 UCSC 浏览器(步骤 1a)中使用的基因组相匹配,然后选择可通过转染载体编码的 Cas9 酶类型识别的 Protospacer Adjacent Motif (PAM)。如果转染载体表达 SpCas9,则选择 20 bp-NGG PAM 格式。单击“提交”以获得针对模板 DNA 的候选 gRNA 序列。b. CRISPOR 算法默认按特异性从高到低对候选 gRNA 序列进行排序,因为这是关键参数。从新页面上出现的候选 gRNA 列表中,选择具有最高麻省理工学院 (MIT) 和切割频率确定 (CFD) 特异性得分的指导序列(Doench 等人,2016 年;Hsu 等人,2013 年;Tycko 等人,2019 年)。这些分数根据以下方面评估候选 gRNA
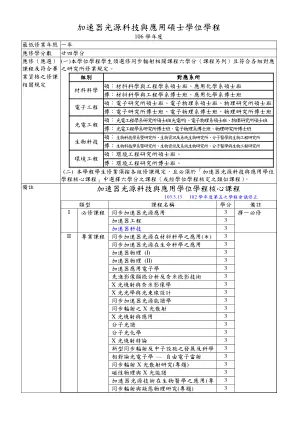
加速器光源科技与应用硕士学位学程
硕士课程在生物学和系统生物学研究所,生物科学与技术系,生物学和系统生物学研究所生物学和系统生物学研究所,生物学科学与技术学院,分子医学与技术研究所,分子医学研究所硕士课程在生物学和系统生物学研究所,生物科学与技术系,生物学和系统生物学研究所生物学和系统生物学研究所,生物学科学与技术学院,分子医学与技术研究所,分子医学研究所

十二个假定的染色体倒位耦合对蜗牛生态型之间的基因流动形成了强大的障碍
染色体重排可导致生殖障碍的耦合,但它们是否以及如何促成物种形成的完成仍不清楚。 Littorina 属的海洋蜗牛反复在分离多个倒位排列的种群之间形成杂交区,为研究它们的屏障效应提供了机会。在这里,我们分析了适应瑞典岛屿不同海浪暴露条件的两种生态型 Littorina fabalis(“大”和“矮”)之间的杂交区中的 2 条相邻横断面。应用全基因组测序,我们发现 17 条染色体中的 9 条有 12 个推定的倒位。其中 9 个推定的倒位在两个生态型之间达到近乎差异固定,并且都处于强烈的连锁不平衡状态。这些倒位覆盖了 20% 的基因组,并携带 93% 的不同单核苷酸多态性 (SNP)。两个横断面的双峰杂交区表明,两种生态型的 Littorina fabalis 在接触后仍保持其遗传和表型完整性。双峰性反映了倒位趋势之间的强耦合和屏障效应在整个基因组中的扩展。人口统计学推断表明,耦合发生在异地时期,并在二次接触后维持了 1,000 代以上。总体而言,这项研究表明,多个染色体倒位的耦合导致了强烈的生殖隔离。值得注意的是,2 个假定的倒位与与近亲物种(Littorina saxatilis)的生态型差异相关的倒置基因组区域重叠,这表明具有相似结构变体的相同区域反复促进了不同物种的生态型进化。

蜡抑制剂PAO85855:倒点抑制剂
PAO85855倒点抑制剂是油的抑郁症的有效添加剂。PAO85855倒点抑制剂设计用于蜡通常结晶的点上游的连续应用。在石油系统中连续使用可以清除一些现有的沉积物,但是PAO85855倾角抑制剂不是批处理处理来“脱水”井或表面设施的溶剂。在石蜡晶体通常形成的点上游的点上连续注入PAO85855倒点抑制剂。

民航工程学(荣誉)工学士学位 - PolyU
5.4 学分转移 19 5.5 零科目注册/休学 20 5.6 一般评估规定 20 5.7 评估原则 21 5.8 评估方法 21 5.9 升学/留校察看/取消注册 22 5.10 重修科目 23 5.11 对评估结果提出上诉/对考试委员会的取消注册决定 23 5.12 特殊情况 24 5.13 评分 25 5.14 不同类型的 GPA 26 5.15 大学毕业要求 27 5.16 奖励分类指南 33 5.17 奖励分类 34 5.18 在学生记录中记录纪律处分 35 5.19 毕业 35 6. 科目大纲 36最终课程文件适用于 2016/17 学年入学学生。课程主办方可随时决定对其进行审查和更改。学生将在适当的时候被告知更改。ii
