XiaoMi-AI文件搜索系统
World File Search System内共生

海洋微生物群中的共生 - 南加州大学多恩西夫分校
共生作用广泛存在于地球生态系统,包括海洋环境。“共同生活”描述了一系列相互作用,从捕食和寄生到共生和互利的正相互作用,这些相互作用通常与共生一词联系在一起。海洋环境中许多众所周知的共生关系涉及微生物与珊瑚等多细胞生物之间的关联,但数十年甚至几个世纪以来,人们一直在通过微观观察描述微生物与微生物之间的共生相互作用。微生物与微生物共生关系的研究一直具有挑战性,部分原因是它们规模小,我们无法在实验室中建立和培养它们,以及用于研究宏观物种的方法无效或不合适。然而,核酸测序、生物信息学、同位素方法和成像方面的技术进步已经开始为这些多样而丰富的相互作用提供新的见解。不依赖培养的方法的应用表明,海洋微生物群落中的微生物相互作用范围从自由生活的浮游细胞之间的代谢物交换到连接共生-细胞器转变的外生和细胞内内共生相互作用。在这里,我们简要概述了共生,然后重点介绍了海洋浮游生物中的两个具体案例——N2-固定和浮游根共生——它们说明了

细菌内生菌来源,概况和动态 - 概念框架
目前,似乎不可想象是对微生物在人类健康或内共生膜中的昆虫中的主要作用不可思议。尽管很久以前发现了微生物内生菌,但对植物相关微生物的作用知之甚少。某些内生菌是水平传播的,而另一些则是种子传播。它们共同影响植物健康。有益的内生菌可以通过增加植物对生物和非生物胁迫的耐药性来促进植物的生长和产量。最近,可用于研究植物生物组的工具有了很大的改进,可以更好地理解在植物一级发生的迷人相互作用。本评论重新定义了“内生菌”和“内生菌”的概念框架,重点是细菌内生细菌的复杂动力学。系统地检查了形成途径和分析内生菌,可以全面探索有关植物 - 微生物相互作用的复杂动力学。此外,对生物和非生物因素如何影响内生细胞组的评估还提供了对植物相关微生物的适应性和弹性的重要见解。我们的综合分析将基因组见解与环境因素相结合,从而对细菌内生菌的功能作用提供了细微的观点。因此,一个新的包容性定义对于准确地表示植物微生物组中相互作用的复杂性以及相关概念的整体情况至关重要。

微生物 - LSTM在线存档
摘要:微生物组组成与疟疾载体中的杀虫剂抗性有关。然而,主要共生体对日益报告的抗药性升级的贡献尚不清楚。这项研究探讨了特定的内共生体Asaia spp。的可能关联,其拟甲虫素耐药性升高,由细胞色素P450S酶和Anopheleles funestus和Anopheles gambiae的电压门控钠通道突变驱动。分子测定法用于检测共生体和电阻标记(CYP6P9A/B,6.5 Kb,L1014F和N1575Y)。总体而言,关键突变的基因分型揭示了与抗性表型的关联。Asaia spp的患病率。在Fumoz_X_FANG菌株中,在5次剂量的Deltamethrin(OR = 25.7; P = 0.002)时与电阻表型相关。蚊子的感染程度更高。此外,丰度与1倍浓度的三分之一的抗性表型相关(p = 0.02,Mann-Whitney测试)。然而,对于Mangoum_x_kisumu菌株,发现的发现却揭示了Asaia载荷与易感表型之间的关联(P = 0.04,Mann-Whitney测试),表明了共生体和苄氯菊酯耐药性之间的负相关。应进一步研究这些细菌,以建立其与其他耐药机制的相互作用,并与其他杀虫剂类别进行交叉抗性。

对帕金森病的影响
摘要:线粒体在调节宿主代谢、免疫和细胞稳态方面发挥着关键作用。值得注意的是,这些细胞器被认为是从 α-变形菌与原始真核宿主细胞或古菌之间的内共生关系进化而来的。这一关键事件决定了人类细胞线粒体与细菌具有一些共同特征,即心磷脂、N-甲酰肽、mtDNA 和转录因子 A,它们可以作为线粒体衍生的损伤相关分子模式 (DAMP)。细胞外细菌对宿主的影响主要通过调节线粒体活动起作用,而且线粒体本身通常就是免疫原性细胞器,可以通过 DAMP 动员触发保护机制。在这项研究中,我们证明暴露于环境中的 α-变形菌的中脑神经元通过 Toll 样受体 4 和 Nod 样受体 3 激活先天免疫。此外,我们还表明中脑神经元会增加与线粒体相互作用的 α-突触核蛋白的表达和聚集,从而导致其功能障碍。线粒体的动态变化也会影响线粒体自噬,这有利于先天免疫信号的正反馈回路。我们的研究结果有助于阐明细菌和神经元线粒体如何相互作用并引发神经元损伤和神经炎症,并使我们能够讨论细菌衍生的病原体相关分子模式 (PAMP) 在帕金森病病因中的作用。

siboglinid小管中微生物氮利用率的钼同位素特征
许多基于化学合成的社区在深海环境中繁荣发展,依赖于硫化物氧化细菌的代谢活性。术后siboglinid tubeworms就是这种情况,其对营养的需求主要通过其endosymbiotic细菌来满足,其中包括在一个称为The Troposomy体的专用器官中。这种化学共生的导致滋养体的氮同位素组成明显低于其他类型的软组织。然而,Sibo Glinids的氮利用的特定过程尚不清楚。作为相关酶(氮酶和硝酸盐还原酶)的关键要素,在氮的生物地球化学循环中是必不可少的。Siboglinids的Mo同位素组成(δ98MO)是解码与氮代谢有关的过程的潜在代理。在这项研究中,我们发现了Δ98mo值沿着南部中国海的Haima渗漏的actimentiferan siboglinid paraescarpia echinospica沿着 - 4.59‰的阴性(-1.13‰±1.75‰±1.75‰±1.75‰,n = 19) - 自然量为Δ98mo的δ98mo值。建议这种极为负的同位素组成是由硝酸盐减少期间的肾小管内共生体或epibionts降低引起的同位素cally light mo引起的。这样的MO同位素签名可以提供一种用于识别Siboglinid Tubeworms的手段,Siboglinid tubeworms是一组因缺乏矿物质骨骼而在岩石记录中由于缺乏矿化骨架而逃脱了明确鉴定的annelids。

从厌氧木质素中分离的 Sodalis ligni 菌株 159R
摘要 具有木质素解聚、分解代谢或两者兼有能力的新型细菌分离物可能与木质纤维素生物燃料应用有关。在本研究中,我们旨在识别能够解决微生物介导的生物技术所面临的经济挑战(例如需要曝气和混合)的厌氧细菌。利用从温带森林土壤中接种并在缺氧条件下以有机溶剂木质素作为唯一碳源进行富集的菌体,我们成功分离出一种新型细菌,命名为 159R。根据 16S rRNA 基因,该分离物属于 Bruguierivoracaceae 科的 Sodalis 属。全基因组测序显示基因组大小为 6.38 Mbp,GC 含量为 55 mol%。为了确定 159R 的系统发育位置,使用 (i) 其最亲属的 16S rRNA 基因、(ii) 100 个基因的多位点序列分析 (MLSA)、(iii) 49 个直系同源群 (COG) 结构域簇和 (iv) 400 个保守蛋白质重建了它的系统发育。分离株 159R 与枯木相关的 Sodalis 行会密切相关,而与采采蝇和其他昆虫内共生体行会关系较弱。估计的基于基因组序列的数字 DNA-DNA 杂交 (dDDH)、基因组保守蛋白质百分比 (POCP) 以及 159R 与 Sodalis 进化枝物种之间的比对分析进一步支持分离株 159R 属于 Sodalis 属的一部分和 Sodalis ligni 的一个菌株。我们建议将之命名为 Sodalis ligni str。 159R (=DSM 110549 = ATCC TSD-177)。

线粒体基因组研究的最新进展
摘要 自从真核生物线粒体中的双链 DNA 被发现以来,它就一直吸引着研究人员,因为它起源于细菌内共生,在编码呼吸复合体亚基方面发挥着关键作用,结构紧凑,并具有特殊的遗传机制。近几年来,高通量测序技术加速了线粒体基因组(线粒体基因组)的测序,并揭示了活体真核生物中组织、基因内容以及复制和转录模式的巨大多样性。一些早期分化的单细胞真核生物谱系保留了某些同源性和基因内容,类似于在 α-变形菌(推测的最接近的线粒体现存群)基因组中观察到的那些,而其他适应厌氧环境的谱系则大幅减少甚至失去了线粒体基因组。在真核生物的三个主要多细胞谱系中,线粒体基因组经历了不同的进化轨迹,其中选择了不同类型的分子(环状与线性、单部分与多部分)、基因结构(有或没有自剪接内含子)、基因内容、基因顺序、遗传密码和转移 RNA 编辑机制。动物进化出了一个相当紧凑的线粒体基因组,长度在 11 到 50 Kb 之间,并且在两侧对称动物中基因内容高度保守,而植物的线粒体基因组较大,为 66 Kb 到 11.3 Mb,具有易于重组的大量基因间重复,真菌的线粒体基因组的大小介于 12 到 236 Kb 之间。
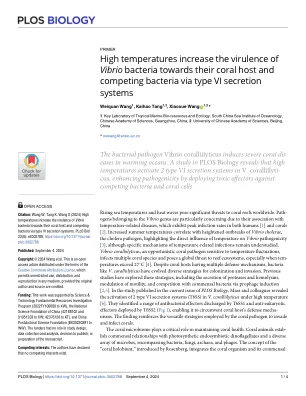
高温增加了通过VI型分泌系统
海上温度和热浪的上升对全世界的珊瑚礁构成了重大威胁。属于弧菌属的途径尤其是由于它们与温度相关疾病的关联,后者在人类和珊瑚[1]和珊瑚[2]中均表现出峰值感染率。夏季温度的升高与霍乱病原体的弧菌病原体的爆发爆发相关,突出了温度对弧菌致病性的直接影响[3],尽管与温度相关感染的特定机制仍然被忽略了。弧菌Coralliilyticus是对温度波动敏感的机会性珊瑚病原体,感染多种珊瑚种类,并对礁生态系统构成全球威胁,尤其是当温度超过27°C时[4]。尽管珊瑚宿主具有多种防御机制,但细菌如V。Coralliilyticus发展了殖民和入侵的多种策略。先前的研究已经探索了这些策略,包括蛋白酶和血素蛋白的分泌,运动能力的调节以及通过预言诱导与共生细菌的竞争[2,5]。在发表在《 PLOS生物学》中发表的研究中,Mass及其同事揭示了V中2型VI型分泌系统(T6SS)的激活。在高温下[6]。他们确定了由T6SS1和T6SS2部署的抗核效应器排放的一系列抗菌效应器(图1),使其能够绕开珊瑚宿主的防御机制。这一发现加强了珊瑚病原体侵入和感染珊瑚的多功能策略。珊瑚微生物组在维持珊瑚健康中起着至关重要的作用。珊瑚动物与光合性内共生鞭毛藻和各种微生物,包括细菌,真菌,古细菌和噬菌体的多种微生物。罗森伯格(Rosenberg

肠道微生物对于昆虫的生存至关重要:它们的起源,功能和进化
抽象的天然生物与周围的物理环境以及广泛的其他生物有密切接触。换句话说,虽然单个生物体构成了生态系统的一部分,但如果它们包括体内存在的各种微生物群落,但它也可以被视为单个生物本身就是建立一个单一的生态系统。 大多数动物都有消化道,喂养,消化,吸收,代谢,排泄和生活。消化道是一个稳定的环境,经常提供丰富的营养,并且居住了微生物。毫不夸张地说,成为动物意味着患有肠道菌群。 微生物的先进材料生产,分解和修饰能力不仅在生态系统中起重要作用,而且在人类社会中也以多种方式使用。特别是,近年来,已经揭示了肠道细菌深深地参与了人类疾病和身体健康,并且细菌在生物体中的多种生物学功能,即共生细菌,引起了人们的关注。 昆虫是人类到目前为止所描述的大多数生物多样性,并且是陆地生态系统的核心生物,但是大多数人都会不断或半稳定地在体内携带微生物。这种现象称为“内部共生”,因为它是一种以无与伦比的空间接近性建立的共生关系,因此观察到了极高的相互作用和依赖性。这些关系通常会创造新的生物学功能。通常,共生的微生物和宿主昆虫几乎彼此融合在一起,形成了一种复合物,好像它是单个生物体一样。同样适用于肠道共生。 共生关系出现了哪些新的生物学功能和现象?通过共同生活,如何将不同生物体的基因组和功能纳入单个生命系统的构建中?共同生活的意义和成本是什么?当个人和个人,自我和非自我融合在一起时会发生什么? 这次,我们将介绍环境适应的演化和机制,可以通过微生物共生,尤其是专注于晚期肠道共生。

线粒体丙酮酸载体抑制剂的最新进展
在真核细胞中,线粒体是内共生器官,与各种细胞过程有关,包括能量消耗,生物合成,信号转移和程序性细胞死亡。1显着,它们是创建三磷酸腺苷(ATP)的主要位置,腺苷三磷酸腺苷(ATP),包括所有生物的通用自由能载体,包括所有五个呼吸链络合物和所有三羧酸周期(TCA)酶。在细胞质和线粒体基质之间的代谢物交换对于执行这些代谢过程是必要的,这些代谢过程仅限于线粒体腔室并保留内部内稳态。电压依赖性阴离子通道允许微小的分子穿过外部线膜。然而,线粒体内膜(IMM)对分子和离子高度渗透,必须依靠特定的转运蛋白和通道来连接细胞质和线粒体的代谢。线粒体载体家族成员执行大部分运输步骤。2其他转运蛋白家族包括线粒体丙酮酸载体(MPC)。3 MPC是一种蛋白质复合物,存在于线粒体内膜中,并负责将丙酮酸从线粒体转运到线粒体基质中,其中丙酮酸转化为乙酰基氧乙烯酶A(乙酰辅酶A)。ace-tyl-coa进入TCA循环,并在其中进一步氧化。另外,线粒体中的丙酮酸也可以通过吡二酸酯羧化酶的羧化来参与糖异生,以产生草乙酸以补充TCA循环。7如上所述,除了被运输到线虫外,丙酮酸还可以通过细胞质中的乳酸脱氢酶(LDH)还原为乳酸。MPC是在1970年代4提出的,最初被称为BRP44L(脑蛋白44样)和BRP44(脑蛋白44)。它在2003年被鉴定在酵母中,并在2012年进一步鉴定在哺乳动物中。3,5,6 MPC是一个相对较小的杂物,由两个亚基组成,分别由12和14 kDa组成,分别为12和14 kDa。