XiaoMi-AI文件搜索系统
World File Search System单倍体

CENH3 错误表达导致柳枝稷单倍体和非整倍体
对于四倍体柳枝稷,我们将单倍体定义为两个亚基因组的基因组拷贝丢失。双单倍体技术需要有效的 2n 诱导系统以及随后的基因组加倍,并将提供新的育种机会,例如为商业杂交生产系统选择高性能自交系。不同柳枝稷亚种群的杂合亲本之间的杂交可产生生物量产量的杂种优势(Bhandari 等人,2017 年;Martinez-Reyna 和 Vogel,2008 年;Vogel 和 Mitchell,2008 年)。然而,由于柳枝稷中活跃的遗传不相容系统以及在获得的相对较少的自交基因型中可能发生的近交衰退和不育,自交系尚未开发。如果有更好的自交系,开发高产单交杂交种将是一种可选的育种方法。由于自交系的性能通常与其杂交种的性能相关,因此选择高产自交系可能具有优势(Hayes & Johnson,1939;Sprague,1977)。此外,DH 技术将促进所需性状、外来基因、转基因、染色体片段或整个染色体的渗入和稳定(Devaux & Pickering,2005;Forster & Thomas,2005)。
圆形染色体会发生减数分裂吗?
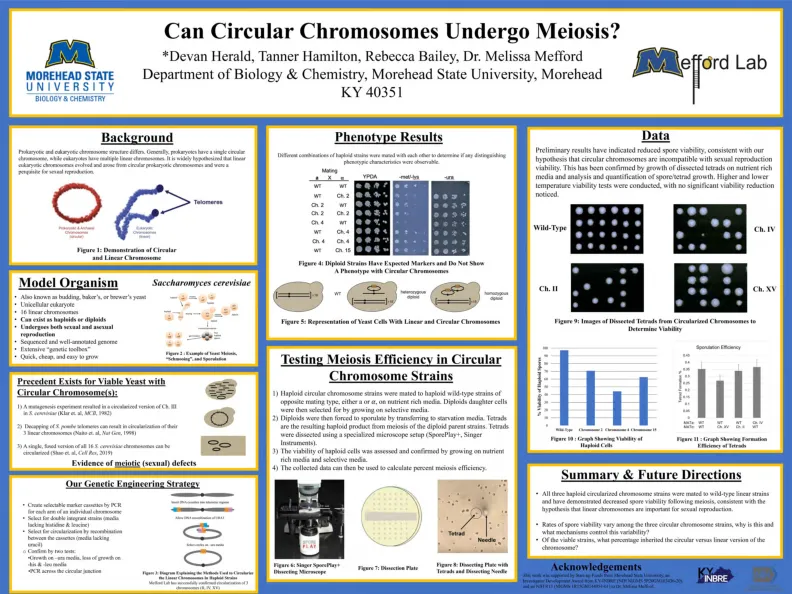
1)单倍体圆形染色体菌株在富含营养培养基上的相反交配类型的单倍体野生型菌株配对。然后,通过在选择性培养基上生长二倍体子细胞。2)然后将二倍体通过转移到饥饿的Me di a来散发。二倍体母体菌株减数分裂是产生的单倍体产物。TETRA DS。3)单倍体细胞的生存能力通过营养素生长

诱导物的父本染色体消除引发油菜双单倍体的诱导
合成的八倍体油菜籽 Y3380 在用作花粉供体为植物授粉时可诱导母本双单倍体。但双单倍体形成的潜在机制仍不清楚。我们推测双单倍体诱导发生在诱导系的染色体传递到母本卵细胞,并通过受精形成合子时。在合子有丝分裂过程中,父本染色体被特异性地消除。在消除过程中,部分父本基因可能通过同源交换渗入母本基因组。然后,合子单倍体基因组加倍(早期单倍体加倍,EH 现象),加倍的合子继续发育成完整的胚胎,最终形成双单倍体后代。为了验证假设,本研究以八倍体Y3380品系为标记,将4122-cp4-EPSPS外源基因回交,得到六倍体Y3380-cp4-EPSPS作为父本材料,对3个不同的母本材料进行授粉。在授粉后48 h观察诱导品系与母本杂交的受精过程,受精率分别达到97.92%和98.72%。授粉12 d后,用原位PCR检测胚中存在cp4-EPSPS,授粉后13 — 23 d,F 1 胚含有cp4-EPSPS基因的概率高达97.27%,而后逐渐下降,在23 — 33 d时为0%。同时免疫荧光观察了3~29天胚胎中cp4-EPSPS的表达情况。随着胚胎的发育,cp4-EPSPS标记基因不断丢失,伴随胚胎死亡,30天后在存活的胚胎中检测不到cp4-EPSPS的存在。同时对诱导后代的SNP检测证实了双单倍体的存在,进一步表明诱导过程是由于父本染色体特异性的丧失引起的。四倍体诱导后代表现出诱导系基因位点的筛选,有杂合性,也有纯合性。结果表明,在诱导过程中,诱导系染色体被消除。

双倍体和单倍体诱导者介导的基因组编辑系统的机会和挑战
摘要:在过去的四十年中,双倍的双倍体在库瑟育种中发挥了重要作用。通过辐照花粉的原位孤立生成是获得单倍倍体的首选技术,然后在葫芦科中将其染色体倍增,例如瓜,黄瓜,南瓜,南瓜和冬南瓜。与其他物种中的单倍体过程加倍相反,库班的原位孤立生成提出了许多限制因素,这些因素阻碍了单倍体的有效产生。此外,这是非常耗时的和劳动力密集的。但是,单倍体诱导者介导的基因组编辑系统是一种可产生双倍双倍体的突破性技术。使用CRISPR / CAS9系统中的几份报告描述了库糖库物种,尽管其应用具有许多瓶颈,但CENH3基因的靶向敲除将允许育种者获得可用于获得多倍性诱导剂线,以获得py源性胚胎。在这篇综述中,我们讨论了使用CURSPR / CAS9技术在葫芦物种中的双倍单倍体和单倍体诱导剂基因型的发展方面取得的进展。本综述为应用单倍体诱导剂介导的基因组编辑系统的应用提供了见解

爸爸不在,但父亲很重要
与大多数真核生物一样,植物携带父本和母本双重基因组。有性生殖允许遗传信息的混合,从而产生多样性,从而可以培育出具有改良农艺性状的新植物品种。然而,植物育种过程通常需要具有固定遗传物质的纯合系或自交系,以评估各种基因组合的性能。在传统育种中生成这些品系是一个耗时的过程,需要多代自交。生产双单倍体植物是获得基因组纯合品系的捷径,只需两代而不是六代或更多代即可实现 1 。玉米育种从这种双单倍体技术中受益匪浅,这要归功于单倍体诱导品系,它可以诱导种子中单倍体胚的形成 1 ( 图 1a )。胚胎发芽成携带一组母体染色体 1 的单倍体幼苗。最近,单倍体诱导系也被巧妙地重新利用,成为将基因组编辑机制引入难以转化的商业作物品种的有力工具 1,2 。尽管是植物育种和研究应用中的有力工具,但植物体内单倍体诱导的分子基础仍然不完整 1 。在本期《自然植物》杂志上,Li 等人 3 发现,突变磷脂酶 D3 基因 (ZmPLD3) 可以诱导母体单倍体胚胎,这为了解这一有趣且有用的生物过程提供了新的思路。

镰状细胞病单倍体骨髓移植国际学习合作 2 期试验
1 医学部血液学/肿瘤学分部和 2 范德堡大学医学院范德堡-梅哈里镰状细胞病卓越中心,田纳西州纳什维尔;3 英国伦敦帝国理工学院圣玛丽医院儿科系;4 荷兰阿姆斯特丹大学阿姆斯特丹大学医学中心血液学系;5 荷兰阿姆斯特丹 Sanquin 研究与兰德斯坦纳实验室血液细胞研究系;6 沙特阿拉伯利雅得费萨尔国王专科医院成人血液学和干细胞治疗及细胞治疗项目;7 巴西圣保罗 Samaritano Higienopolis-Americas 医院儿科造血细胞移植项目; 8 儿科造血干细胞移植项目,儿科造血细胞移植,Instituto de Oncologia Pediatrica – Graacc/Unifesp,圣保罗,巴西; 9 儿科骨髓移植项目,Pequeno Príncipe 医院/Instituto de Pesquisa Pele Pequeno Principe,巴西库里蒂巴; 10 圣保罗大学里贝朗普雷托医学院医学系、11 血液学系和 12 临床肿瘤学系,巴西圣保罗; 13 小儿血液科/肿瘤科,阿卜杜勒阿齐兹国王医学城,国民警卫队卫生部,阿卜杜拉国王国际医学研究中心,沙特本阿卜杜勒阿齐兹国王健康科学大学,沙特阿拉伯利雅得; 14 北卡罗来纳州夏洛特市维克森林大学医学院 Atrium Health Levine 儿童医院儿科、儿科移植和细胞治疗系;15 佛罗里达州盖恩斯维尔市佛罗里达大学儿科、儿科骨髓移植和细胞治疗项目系;16 俄亥俄州克利夫兰市克利夫兰诊所儿科血液学/肿瘤学、儿科血液肿瘤学和血液与骨髓移植系;17 法国巴黎圣路易斯医院血液学、青少年和青年血液学系;18 俄亥俄州哥伦布市全国儿童医院儿科血液学、肿瘤学、血液和骨髓移植系;19 田纳西州纳什维尔市范德比尔特大学医学中心药学服务系; 20 田纳西州纳什维尔范德堡大学医学院儿科、血液学/肿瘤学系;21 伊利诺伊州芝加哥 Rodeghier 咨询公司

在单倍体组织 - 特异性基因的强烈纯化选择支持掩盖理论
掩盖理论指出,单倍体阶段表达的基因将在更有效的选择下。在con trast中,选择在二倍体阶段表达的基因中的效率较低,在二倍体阶段,隐性有害或有益突变的适应性可能以杂合形式隐藏。这种差异可以在流动性中几个进化过程,例如维持遗传变异,适应率和遗传负荷。掩盖理论期望已在单细胞单倍体和二倍体生物中得到证实。然而,在多细胞生物(例如植物)中,单倍体选择的作用并不明确。在植物中,已经使用血管中的雄性单倍体组织进行了大量选择的研究。因此,这些系统中的证据与性选择和种内竞争的影响相混淆。其他植物群的证据很少,结果没有对掩盖理论的支持。在这里,我们使用了裸子苏格兰松树巨型植物学,母体衍生的种子单倍体组织和四个二倍体Tis SU来测试在具有组织特异性表达的一组基因上纯化选择的强度。通过使用这些基因的靶向重新定位数据,我们获得了遗传多样性,0倍和4倍位点的位点频谱的估计值,并推断了单倍体组织和二倍体组织 - 特异性基因中新突变的适应性效应的分布。我们的结果表明,在单倍体巨脂组织组织中表达的组织特异性基因纯化选择更强,并且这种强选择的信号不是由高表达水平驱动的伪像。

通过局部单倍体人类多能干细胞对 BRCA2 基因变体进行功能注释
BRCA2 基因突变与散发性和家族性癌症有关,可导致基因组不稳定并使癌细胞对聚(ADP-核糖)聚合酶 (PARP) 抑制敏感。本文表明,删除一个 BRCA2 拷贝的人类多能干细胞 (hPSC) 可用于注释此基因的变体并测试其对 PARP 抑制的敏感性。通过使用 Cas9 编辑局部单倍体 hPSC 和从其分化的成纤维细胞中的功能性 BRCA2 等位基因,我们鉴定了该基因中的必需区域以识别允许突变和功能丧失突变。我们还使用 Cas9 直接测试单个氨基酸的功能,包括由意义不明确的临床 BRCA2 变体编码的氨基酸,并鉴定了对用作 BRCA2 缺陷型癌症治疗标准的 PARP 抑制剂敏感的等位基因。局部单倍体人类多能干细胞可以促进基因的详细结构功能分析以及临床观察到的突变的快速功能评估。

RNAi介导的AccenH3的下调可以在洋葱中诱导体内单倍体(Allium cepa L.)
作为男性父母,事件E1,E2和E5的相对种子集效率分别为37.89%,61.82%和83.76%(表1和补充图。9)。这些发现进一步表明,Accenh3的敲低影响了种子集。差异种子集可能是在相互交叉中观察到的转基因偏置隔离变形的原因之一。我们的观察结果

通过标记辅助QHIR1和QHIR8在玉米中加速单倍诱导率和单倍体验证
双倍(DH)技术更常规地应用于玉米杂种繁殖中。但是,单倍诱导和识别的某些问题持续存在,需要解决以优化DH生产。我们的目标是使用taqman测定法实施QHIR1(MTL/ ZMPLA1/ NLD)和QHIR8(ZMDMP)的同时进行标记辅助选择(MAS),以在F 2代生成四个BHI306衍生的热带热带×温度诱导剂中。我们还旨在评估F 3代的单倍体诱导率(HIR)作为对MAS的表型反应。我们强调了每个诱导剂家族的HIR的显着增加。携带QHIR1和QHIR8的基因型比仅携带QHIR1的基因型表现出1-3倍的单倍体频率。此外,QHIR1标记还用于在种植后7天验证推定的单倍体幼苗。流式细胞仪分析是评估R1-NJ和QHIR1标记的准确性的黄金标准测试。QHIR1标记显示出很高的精度,并且可以在早期幼苗阶段通过R1-NJ标记在早期幼苗阶段进行多个单倍体识别。