XiaoMi-AI文件搜索系统
World File Search System合酶

Primestar®LongSeqDNA聚合酶
请注意,此产品仅用于研究用途。它不打算用于人类或动物的治疗或诊断程序。另外,请勿将此产品用作食品,化妆品或家居用品等。takara产品不得转售或转让,修改用于转售或转让,或无需未经Takara Bio Inc.的书面批准而用于制造商业产品。如果您需要其他使用许可证,请通过我们的网站www.takarabio.com与我们联系。您对此产品的使用也符合产品网页上所述的任何适用许可要求。您有责任审查,理解并遵守此类陈述所施加的任何限制。所有商标都是其各自所有者的财产。某些商标可能不会在所有司法管辖区注册。

Pfu DNA 聚合酶综合综述
Pfu DNA 聚合酶是一种源自超嗜热古菌 Pyrococcus furiosus 的耐热酶,因其高保真度和强大的加工性而广受认可。它的 3'-5' 核酸外切酶活性使其成为正确扩增短链和复杂 DNA 链不可或缺的酶。Pfu DNA 聚合酶的这些生化特性促进了其提取和生产方法的重大进步。本综述涵盖了一些传统的纯化方法,包括蛋白质纯化和亲和层析,以及重组基因表达、自动化生产系统和基于膜的技术的最新进展。最近开发了新的酶工程方法,例如 CRISPR-Cas9 介导的基因优化,这提高了提取效率的标准以满足新兴需求。曾经具有挑战性的 Pfu DNA 聚合酶生产已通过在实验室和商业规模的大肠杆菌中重组表达得到了显着简化。涉及 IPTG 浓度和响应面方法的优化技术已将产量提高了 30%。自诱导意味着可以实现更高的生物量输出。如今,Pfu DNA 聚合酶的应用范围从标准 PCR 到分子生物学、法医分析、临床微生物学和生物技术领域的高级临床诊断。
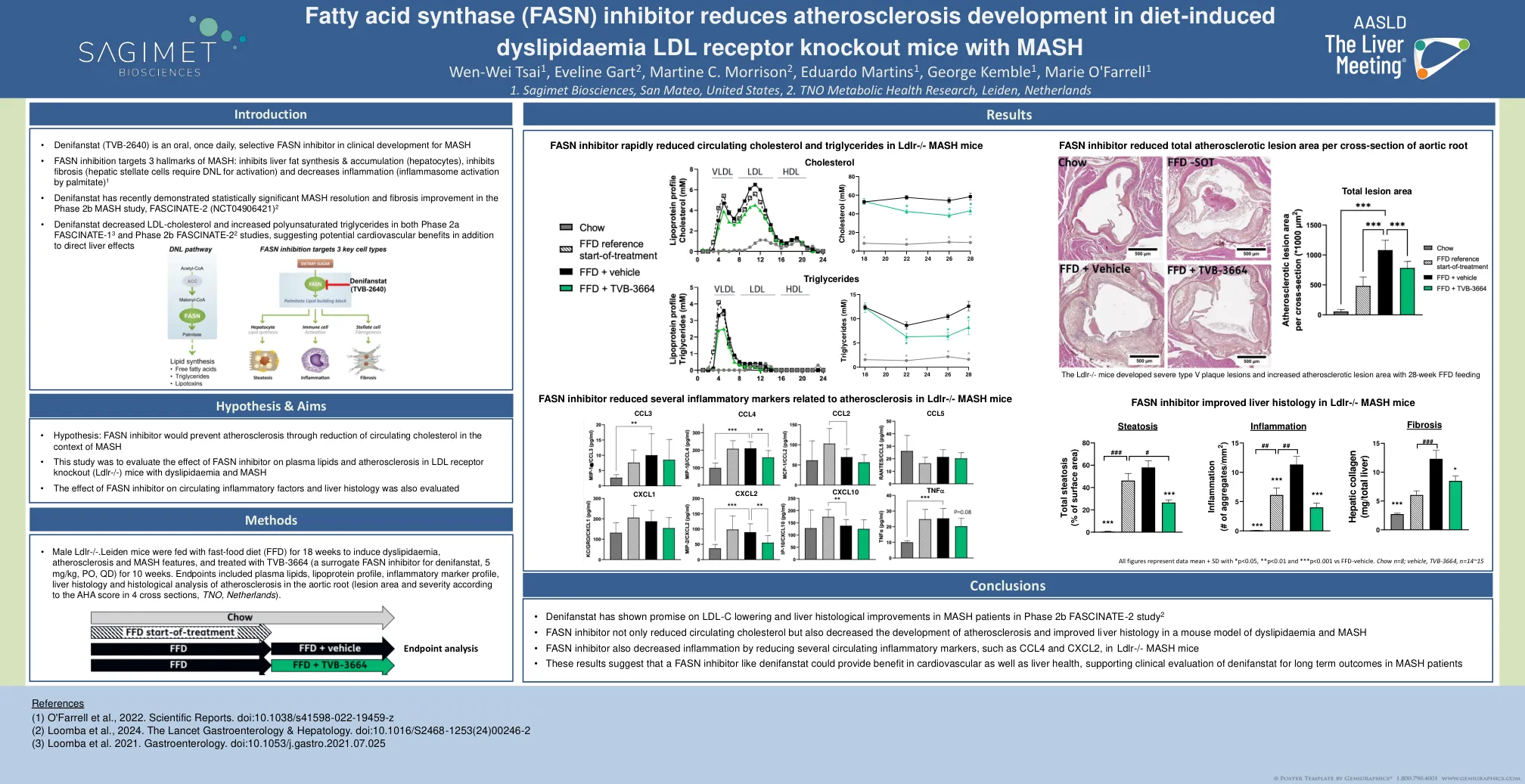
脂肪酸合酶 (FASN) 抑制剂可减少饮食引起的动脉粥样硬化的发展
• 雄性 Ldlr-/-.Leiden 小鼠以快餐饮食 (FFD) 喂养 18 周以诱发血脂异常、动脉粥样硬化和 MASH 特征,并用 TVB-3664(denifanstat 的替代 FASN 抑制剂,5 mg/kg,PO,QD)治疗 10 周。终点包括血浆脂质、脂蛋白谱、炎症标志物谱、肝脏组织学和主动脉根部动脉粥样硬化的组织学分析(根据 4 个横断面的 AHA 评分确定病变面积和严重程度,荷兰 TNO)。

棉花中的δ-杜松烯合酶基因家族
棉酚是棉花 ( Gossypium hirsutum L.) 中常见的一种萜醛,对植物抵御害虫和病原体至关重要。然而,其固有毒性限制了棉籽在食品和饲料中的使用。这项研究重点验证了 (+)-delta- 杜松烯合酶基因家族的表达模式,该基因家族在棉酚的生物合成中起着至关重要的作用。我们的目标是利用这些信息指导基因组编辑策略,以降低棉籽中的棉酚水平。我们使用定量实时 PCR (qRT-PCR) 分析了 32 个 (+)-delta-杜松烯合酶基因在胚珠和叶片中的表达,涵盖六个发育阶段,从开花后 (DPA) 20 到 45 天,每隔五天一次。我们的结果显示,无论处于哪个发育阶段,都有 10 个基因在胚珠中表达。其中,六个基因:Gohir.A04G023700、Gohir.D05G363800、Gohir.A08G087000、Gohir.D05G363900、Gohir.D05G364000 和 Gohir.D05G364300,在各个阶段始终表现出明显更高的表达水平。值得注意的是,Gohir.D05G363900、Gohir.D05G364000 和 Gohir.D05G364300 在所有阶段都表现出略高的表达水平,使其成为靶向基因组编辑的合适候选基因。这些发现为 (+)-delta- cadinene 合酶基因家族的表达动态提供了宝贵的见解,并确定了未来基因组编辑实验的潜在靶基因,旨在通过降低棉酚含量来提高棉籽的利用率。

使用 Qiagen HotStar Taq 聚合酶试剂盒进行 DNA PCR 检测
注意:主混合物应在指定的“洁净室”区域制备,并且切勿将提取液/模板带入“洁净室”。主混合物制备完成后,在制备主混合物的洁净罩中,或在打开 DNA/RNA 模板(脏)罩中的 DNA 模板(提取的样本)之前,向每个 PCR 反应管中添加 20 或 45 或 90μl 主混合物。

phi29 DNA聚合酶
注意 本产品仅供研究使用。它不适用于人类或动物的治疗或诊断程序。此外,请勿将本产品用作食品、化妆品或家居用品等。未经 Takara Bio Inc. 书面批准,不得转售或转让、修改以进行转售或转让或用于制造商业产品。如果您需要其他用途的许可,请通过我们的网站 www.takarabio.com 与我们联系。您对本产品的使用还需遵守产品网页上描述的任何适用许可要求。您有责任查看、理解并遵守此类声明所施加的任何限制。所有商标均为其各自所有者的财产。某些商标可能并未在所有司法管辖区注册。

脂肪酸合酶 2 敲低改变了飞蝗感染藤黄微球菌期间免疫和生殖之间的能量分配策略
免疫与生殖是雌性昆虫生存和种群维持的重要功能。然而由于资源有限,这两个功能无法同时满足,从而导致它们之间需要进行能量权衡。值得注意的是,这种免疫-生殖权衡的机制尚不清楚,而能量竞争可能在其中起着核心作用。本研究以飞蝗为研究对象,对参与脂质合成和昆虫能量代谢的关键基因脂肪酸合酶(FAS)进行了研究。利用细菌感染和RNA干扰(RNAi)技术研究了不同处理下蝗虫的免疫、繁殖力和能量代谢模式的变化。本研究结果表明,藤黄微球菌感染可触发蝗虫的免疫反应,显著上调防御素3(DEF3)和Attacin的表达,并增强酚氧化酶(PO)活性。当 FAS2 沉默后,细菌攻击在较小程度上上调了 DEF3 和 Attacin 的表达,导致溶菌酶活性增加而不是 PO。此外,细菌感染导致脂肪体中糖原和葡萄糖含量降低,同时三酰甘油(TAG)含量显著增加。然而,在 FAS2 敲低后,脂肪体中的脂质和碳水化合物含量均显著降低。与单独的细菌感染相比,低 FAS2 表达进一步加剧了蝗虫的繁殖力受损。卵黄蛋白 A ( VgA ) 和卵黄蛋白 B ( VgB ) 的表达水平显著降低,卵巢萎缩严重。值得注意的是,卵巢重量仅为对照组的 21%。此外,雌性表现出最少的产卵行为。总之,我们的研究结果表明,在 FAS2 基因沉默后,蝗虫更倾向于免疫刺激能量激活,而生殖投入减少。该研究成果将有助于进一步探索蝗虫免疫和生殖能量之间权衡的分子机制。

使用化学和遗传方法评估利什曼原虫肌醇磷酸神经酰胺合酶作为药物靶点
摘要:有效疫苗的缺乏和对当前治疗方法的耐药性的产生凸显了对新型抗利什曼原虫药物的迫切需求。鞘脂代谢被认为是利什曼原虫特异性靶点的有希望的来源,因为这些脂质是真核生物质膜的关键结构成分,并参与不同的细胞事件。肌醇磷酸神经酰胺 (IPC) 是利什曼原虫中的主要鞘脂,是 IPC 合酶 (IPCS) 介导的反应的产物。抗组胺药富马酸氯马斯汀已被确定为 L. major 中的 IPCS 抑制剂和体内强效的抗利什曼原虫。在这里,我们试图进一步研究这种化合物在更易处理的物种 L. mexicana 中的靶点,采用结合基因组学、蛋白质组学、代谢组学和脂质组学技术以及分子和生化研究的方法。虽然数据表明对富马酸氯马斯汀的反应基本保持不变,但发现了鞘脂代谢以外的意外干扰。此外,虽然删除编码 Lmx IPCS 的基因在体外影响不大,但它确实影响了富马酸氯马斯汀的疗效,更重要的是,影响了体内致病性。总之,这些数据表明氯马斯汀确实抑制了 Lmx IPCS 并导致相关的代谢紊乱,但其主要目标可能在其他地方。关键词:利什曼原虫、肌醇磷酸神经酰胺合酶、富马酸氯马斯汀、多组学、CRISPR-Cas9、热蛋白质组学分析

重组人类DNA指导的DNA/RNA聚合酶mu(POLM)
序列 MLPKRRRARVGSPSGDAASSTPPSTRFPGVAIYLVEPRMGRSRRAFLTGLAR SKGFRVLDACSSEATHVVMEETSAEEAVSWQERRMAAAPPGCTPPALLDISW LTESLGAGQPVPVECRHRLEVAGPRKGPLSPAWMPAYACQRPTPLTHHNTGL SEALEILAEAAGFEGSEGRLLTFCRAASVLKALPSPVTTLSQLQGLPHFGEHSS RVVQELLEHGVCEEVERVRRSERYQTMKLFTQIFGVGVKTADRWYREGLRTL DDLREQPQKLTQQQKAGLQHHQDLSTPVLRSDVDALQQVVEEAVGQALPGA TVTLTGGFRRGKLQGHDVDFLITHPKEGQEAGLLPRVMCRLQDQGLILYHQH QHSCCESPTRLAQQSHMDAFERSFCIFRLPQPPGAAVGGSTRPCPSWKAVR VDLVVAPVSQFPFALLGWTGSKLFQRELRRFSRKEKGLWLNSHGLFDPEQKT FFQAASEEDIFRHLGLEYLPPEQRNA

重组 Cenarchaeum symbiosum DNA 聚合酶 II 大亚基 (polC),部分
Cenarchaeum symbiosum DNA 聚合酶 II 大亚基 (polC) 是古菌 Cenarchaeum symbiosum 中 DNA 复制过程中的关键酶。其主要功能包括在细胞分裂和基因组维持过程中合成 DNA 链。其研究领域包括揭示其在基因组稳定性中的作用以及探索其在研究古细菌遗传学中的应用。在分子生物学和古细菌遗传学中,polC 是理解 DNA 复制机制的关键焦点。该酶的重要性在于提供对基本生物过程的洞察,促进古细菌分子生物学的进步。