XiaoMi-AI文件搜索系统
World File Search System基因组


基因组编辑技术及未来发展
[1] Kim YG, Cha J, Chandrasegaran S. 混合限制性内切酶:锌指融合至 Fok I 切割域。美国国家科学院院刊,1996,93:1156-60 [2] Boch J, Scholze H, Schornack S 等人。破解 TAL 型 III 效应物的 DNA 结合特异性密码。科学,2009,326:1509-12 [3] Moscou MJ, Bogdanove AJ。一个简单的密码控制 TAL 效应物的 DNA 识别。科学,2009,326:1501 [4] Jinek M, Chylinski K, Fonfara I 等人。适应性细菌免疫中的可编程双 RNA 引导 DNA 内切酶。 Science, 2012, 337:816-21 [5] Wyman C, Kanaar R. DNA双链断裂修复:结局好一切都好。Annu Rev Genet, 2006, 40:363-83 [6] Komor AC, Kim YB, Packer MS等人。无需双链DNA切割即可对基因组DNA中的目标碱基进行可编程编辑。Nature, 2016, 533:420-4 [7] Nishida K, Arazoe T, Yachie N等人。利用混合原核和脊椎动物适应性免疫系统进行靶向核苷酸编辑。Science, 2016, 353:8729 [8] Gaudelli NM, Komor AC, Rees HA等人。无需DNA切割即可对基因组DNA中的A*T进行可编程碱基编辑为G*C。 Nature, 2017, 551: 464-71 [9] Kurt IC, Zhou R, Iyer S, et al. CRISPR C-to-G 碱基编辑器用于诱导人类细胞中的靶向 DNA 颠换。Nat Biotechnol, 2021, 39: 41-6 [10] Zhao D, Li J, Li S, et al. 糖基化酶碱基编辑器可实现 C-to-A 和 C-to-G 碱基变化。Nat Biotechnol, 2021, 39: 35-40 [11] Anzalone AV, Randolph PB, Davis JR, et al. 搜索和

人原代T细胞基因组编辑
( A )使用ImmunoCult™ 人 CD3 / CD28 或 CD3 / CD28 / CD2 T 细胞激活活化剂人 T 2 - 3 天后,通过将 TCR αβ 和 CD3 受体与抗体结合,进行流式分析,来测定 TRAC 的敲除效率。每个条件的每个数据点代表一个单独的供体;n = 4 - 8 个供体。每一列线路表示干±标准差。( B ) )首先人T细胞被ImmunoCult™人CD3 / CD28 T细胞剂激活活化剂3天,然后进行电转。在电转48小时后,通过ArciTect™ T7循环内切酶I试剂盒测定基因组编辑(切割)的效率。 RNP 电转:+ RNP 。( C - D )被ImmunoCult™ 人 CD3 / CD28 T 细细胞激活剂活化 3 天的人 T 细胞经( C )模拟电转(无 RNP )和( D ) RNP 电转后 TCR αβ 和 CD3 的流式分析点图。( E )被ImmunoCult™ 人 CD3 / CD28 T 细胞激活剂活化 3 天的人 T细胞的CD4和CD8流式分析点图。

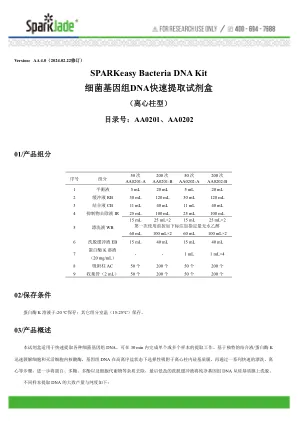
SPARKeasy Bacteria DNA Kit 细菌基因组DNA快速 ...
▲ 可选步骤:如果 RNA 残留较多,需要去除 RNA ,可以在加入 200 μL 结合液 CB 前加 4 μL RNase A ( 100 mg/mL )溶液,振荡混匀,室温放置 5-10 min 。

HPLC技术对细菌基因组DNA碱基组成的精确测定
'e kn---- adenkn〜Cal----甘尼:cn------------------------- i jrdcl【:5 r l〜l〜l〜oslno l〜oslno l〜oslno;)

非洲绿猴(African Green Monkey)全基因组CRISPR sgRNA ...
目的基因 sgRNA 数目: 64853 ;阴性对照 sgRNA 数目: 2000 ; sgRNA 大小: 20bp


基因组技术和基因组医学
12:35-13:00对肌肉蛋白基因在肥厚型心肌病的发展中的影响,使用诱导的多能干细胞技术和CRISPR/CAS9编辑Elena Dementyeva博士,开发镜学研究所的实验室高级科学家Elena Dementyeva博士,细胞遗传学研究所,遗传学和基因学研究所,基因学SB。
