XiaoMi-AI文件搜索系统
World File Search System增强子

新辅助治疗HER2-阳性乳腺癌中的HER2靶向疗法:系统评价和网络元分析Gunasekara,A。D. M
摘要:在三十多年来,基于肿瘤选择性治疗实体瘤的渗透性和保留率(EPR)效应的纳米医学已受到了很大的关注。然而,由于肿瘤或栓塞性肿瘤血管,晚期癌症的治疗仍然是一个巨大的挑战,这导致了EPR效应的所谓异质性。我们先前使用一氧化氮供体和其他称为EPR效应增强子的药物来恢复血管中血管中血流受损的方法。在这里,我们表明,两个新型的EPR效应增强剂 - 异端二硝酸盐(ISDN,Nitrol®)和Sildena fi柠檬酸盐 - 将三种大分子分子药物递送至肿瘤:聚(造型(造型(造型))(造型 - co-maleic Acid)(Sma)和cisplatin,smaplatin,smaplatin,smaplatin;聚(N-(2-羟丙基)甲基丙烯酰胺)聚合物共轭锌原磷脂(光动力疗法和成像);和SMA葡萄糖胺 - 偶联的硼酸络合物(硼中子捕获疗法)。我们在患有晚期C26肿瘤的小鼠中测试了这些纳米果。当这些纳米医学与ISDN或Sildena-Fil一起施用时,肿瘤递送,因此阳性治疗结果在直径为15 mm或更多的肿瘤中增加了2至4倍。这些结果证实了使用EPR效应增强子恢复肿瘤血流的基本原理。总而言之,所有测试的EPR效应增强剂均显示出在癌症治疗中应用的巨大潜力。
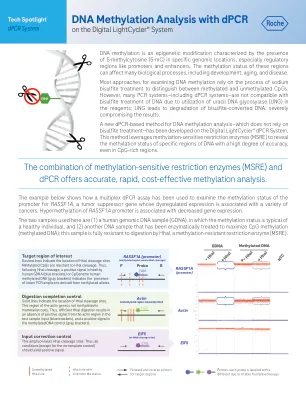
DNA甲基化分析DPCRDNA甲基化分析DPCR
高-GC区域(例如RASSF1A启动子)的扩增通常效率低下,即使在未甲基化的区域中也是如此(请参阅下图中图中的第一组数据;测试的GC的GC含量为72%GC)。这可能会使不论甲基化状态如何准确量化模板的数量。然而,在使用DigitalLightCycler®DPCR系统时,将高GC增强子添加到扩增反应中可以显着提高放大效率和定量准确性(请参见下文);这增加了基于DPCR的DNA甲基化分析的灵敏度和准确性。

使用深度学习的建模胰岛增强剂可以确定与T2D和血糖性状相关的基因座的候选因果变异。
遗传关联研究已经确定了数百个与2型糖尿病(T2D)和相关性状相关的独立信号。尽管取得了这些成功,但鉴定遗传关联信号基础的特定因果变异仍然具有挑战性。在这项研究中,我们描述了一种深度学习(DL)方法,以分析序列变体对增强子的影响。专注于胰岛(T2D相关组织),我们表明我们的模型学习了胰岛特异性转录因子(TF)调节模式,可用于优先考虑候选因果变体。在与T2D和相关血糖性状相关的101个遗传信号中,在链接不平衡中发生多种变体,我们的方法提名每个关联信号的单个因果变体,包括先前显示的三种变体在胰岛含量与含量的细胞类型中改变了报告基因的活性。对于与血糖水平相关的另一个信号,我们使用胰岛β细胞系中的统计细胞映射测试所有候选因果变异,并显示出对模型定位变体TF结合的等位基因影响的生化证据。为了帮助未来的研究,我们公开分发了约6700万个遗传变异的模型和胰岛增强子扰动分数。我们预计,本研究中提出的DL方法将增强候选因果变异的优先级,用于功能研究。

激发教育和 - 内部salk
胰腺癌是最具侵略性,致命的肿瘤类型之一,多年来,研究人员一直在努力开发针对肿瘤的有效药物。现在,第一作者Corina Antal和同事Ronald Evans教授已经确定了一套新的分子,这些分子为胰腺癌腺癌(PDAC)(最常见的胰腺癌类型)增添了胰腺导管腺癌(PDAC)的生长。新的研究解释了某些基因突变如何通过激活打开其他基因的“超级增强子”来引发胰腺癌的控制外增长。他们还显示了一种新药的有效性,该药物通过阻止了超级增强剂的影响,从而使胰腺癌生长刹车。

糖皮质激素诱导增强剂的通量筛选......
压力暴露与多种精神疾病风险增加有关。压力对个体的长期影响可能受到遗传变异的影响。使用大规模并行增强子分析,我们确定了位于基因调控元件中的遗传变异,这些元件调节对糖皮质激素(一种主要应激激素)的转录组反应。这些变异富含与精神疾病和神经行为特征相关的变异,并影响精神疾病患者死后大脑中发生改变的转录本。此外,这些遗传变异与生理应激反应的变化有关。总之,这表明这些功能性遗传变异可能与精神病理学有因果关系,可能是通过影响生理应激反应和应激反应性大脑转录来实现的。

Miodrag Grbic 的基因组学技术创造了...
每个生物体的基因组都代表着一组蛋白质编码基因、调控非编码 RNA 基因和调控元件(启动子和增强子),它们指导着生物体的发育。DNA 中编码的这些信息对于生物体的生长和繁殖、潜在的生理过程、行为和整体表型至关重要。此外,DNA 序列还包含有关进化历史、疾病风险因素的信息,并决定对外部环境的反应。随着大规模测序技术的发展,我们可以从基因组数据中提取大量信息。因此,基因组工具在几乎所有研究领域、个性化医疗、疾病预防、农业和新技术开发中都变得至关重要。

设计师基因由人工智能提供
核心启动子不仅确定基因词的启动何方,还决定基础和增强子诱导的条件下的转录活性。在核心启动子内的多个短序列元素已在不同的物种中鉴定出来,但是它们如何共同发挥作用以及它们在何种程度上是真正物种特异性的。在本期的基因和发展中377 - 382)报告合成核心启动子的大规模平行测量值,以生成大量的激活数据集,以告知统计学习模型以识别人和果蝇核心启动子的序列差异。然后将此机器学习模型应用于设计基因核心启动子,这些基因核心启动子特别针对人类转录机械。

MYT1L 是抑制成年小鼠大脑早期神经元发育程序所必需的
体外研究表明,神经发育障碍基因髓鞘转录因子 1 样 (MYT1L) 在成纤维细胞向神经元直接分化过程中抑制非神经元谱系基因。然而,MYT1L 在成年哺乳动物大脑中的分子和细胞功能尚未完全确定。在这里,我们发现 MYT1L 的缺失会导致深层 (DL) 基因表达上调,这对应于成年小鼠皮质中 DL/UL 神经元的比率增加。为了确定潜在的机制,我们进行了靶向切割和使用核酸酶释放 (CUT&RUN) 以绘制 MYT1L 结合靶标和 MYT1L 缺失后小鼠发育皮质和成人前额叶皮质 (PFC) 中的表观遗传变化。我们发现 MYT1L 主要与开放染色质结合,但启动子和增强子之间具有不同的转录因子共占。同样,多组学数据集整合表明,在启动子处,MYT1L 的缺失不会改变染色质的可及性,但会增加 H3K4me3 和 H3K27ac,从而激活一组早期神经元发育基因以及 Bcl11b(DL 神经元发育的关键调节因子)。同时,我们发现 MYT1L 通常通过关闭染色质结构和促进活性组蛋白标记的去除来抑制与神经元迁移和神经元投射发育相关的神经源性增强子的活性。此外,我们还表明 MYT1L 在体内与 HDAC2 和转录抑制因子 SIN3B 相互作用,这为抑制组蛋白乙酰化和基因表达提供了潜在机制。总体而言,我们的研究结果提供了 MYT1L 体内结合的全面图谱,并提供了有关 MYT1L 缺失如何导致成年小鼠大脑中早期神经元发育程序异常激活的机制见解。

保守元素中人类特异性缺失的功能和进化影响
结果:我们通过检查各种脊椎动物基因组的保守区域并与自信注释的人类特异性固定缺失重叠,从而确定了10,032个HCONDEL。我们发现,这些HCONDEL富含源自茎羊膜的保守序列。与转录,表观基因组和表型数据集重叠均暗示神经元和认知功能影响。我们在六种不同的人类细胞类型中使用MPRA表征了这些HCONDEL,包括诱导多能干细胞衍生的神经祖细胞。我们发现800个HCONDEL显示出物种特异性的调节作用。尽管许多HCONDELS扰动转录因子 - 有效增强子中的结合位点,但我们估计30%创建或改善了结合位点,包括激活剂和阻遏物。

基因组学和人类遗传学的年度综述自然和基因调节区域的实验重新布线
复杂生物的成功发展和持续的功能取决于遗传密码的忠实执行。此过程中的关键步骤是基因的正确空间和时间表达。基因精心策划的转录主要由顺式调节元素控制:启动子,增强子和绝缘子。可以通过改变顺式调节元件的变形和遗传变体导致单基因和复杂性疾病和癌症的频率来看出这种关键生物学过程的医学重要性。在这里,我们提供了可用于表征和扰动基因调节电路的方法的概述。然后,我们强调了调节性重新布线有助于疾病的机制,并以我们如何使用我们对基因调节的理解来改善人类健康的观点。