XiaoMi-AI文件搜索系统
World File Search System大鼠

大鼠 Chiari 样畸形的手术建模
在人类医学中, Chiari 畸形包括一组影响后颅窝和颅颈交界处的多种畸形。 这些畸形的特征是小脑扁桃体突出于枕骨大孔下方,有时伴有不同程度的下降到脑干 [ 1 ]。 与 Chiari 1 型类似,在小型犬中发现的最常见的 Chiari 畸形类型被国际兽医工作组命名为 Chiari 样畸形 (CLM) [ 1 ]。 由于颅骨和颅颈交界处的形态畸形导致 CLM 的尾颅窝 (CCF) 相对较小,导致 CCF 体积与脑实质不一致 [ 2 , 3 ]。这种差异造成了 CCF 拥挤,使小脑通过枕骨大孔向尾部移位 [2-4]。由于颅骨功能不全导致的小脑疝与继发性脊髓空洞症 (SM) 有关,因为它会改变正常的脑脊液 (CSF) 流动,而这种改变是由于背部颅颈蛛网膜下腔阻塞和脊髓扭结所致 [5, 6]。磁共振成像 (MRI) 是 CLM 诊断的金标准,它通过 T1 加权 (T1W) 和 T2 加权 (T2W) 矢状面和横向成像来评估大脑和颈部的线性和体积变化 [2, 6]。矢状面 T2W 图像是最有价值的序列 [5]。诊断是通过识别异常发现来进行的,例如小脑尾部疝、枕骨发育不良导致的小脑尾部受压以及脑脊液衰减 [ 2 , 5 ]。由于缺乏明确的病理机制和治疗方法,Chiari 畸形对人类和兽医研究提出了重大挑战。此外,缺乏通过获得性手段在实验动物中诱发 CLM 的方法。特别是据我们所知,目前还没有已知的诱发 CLM 的方法。由于缺乏适合实验研究的动物模型,目前的状况更加糟糕,阻碍了对 Chiari 畸形病理机制的深入了解并阻碍了有效治疗策略的制定。因此,本研究旨在通过介绍大鼠 CLM 模型的开发来解决这一知识空白,该模型可作为推进未来 Chiari 病研究的基本工具。具体而言,本研究通过使用手术方法提出了一种可靠且新颖的方法来创建第一个 CLM 模型。

大鼠轻度爆炸性脑损伤后的社会心理损害
† 通讯作者 Riyi Shi,医学博士,哲学博士,625 Harrison St,West Lafayette,IN 47907,电话:(765)-494-6446,传真:(765)-494-7605,riyi@purdue.edu,William A. Truitt,哲学博士,320 W 15th St Office #314G,Indianapolis,IN 46202,电话:(317)-278-9050,btruitt@iu.edu。 * 同等贡献作者 作者贡献(CRediT 声明) Nicholas Race:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Katharine Andrews:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Elizabeth Lungwitz:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Sasha Vega-Alvarez:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Timothy Warner:调查、分析、写作 - 修订 Glen Acosta:调查、分析、写作 - 修订 Jiayue Cao:调查、分析、写作 - 修订 Kun-han Lu:调查、分析、写作 - 修订 Zhongming Liu:概念化、方法论、分析、写作 - 修订、监督 Amy Dietrich:调查、验证、分析、写作 - 修订 Sreeparna Majumdar:调查、验证、分析、写作-修改 Anantha Shekhar:分析、写作-修改、监督 William Truitt:概念化、方法论、分析、写作-修改、监督 Riyi Shi:概念化、方法论、分析、写作-修改、监督

酪氨酸羟化酶 TH-Cre 敲入大鼠
该模型在内源性酪氨酸羟化酶 (TH) 启动子的控制下表达 cre-重组酶,从而能够在多巴胺能神经元中进行特异性表达。该模型在 TH 开放阅读框的翻译终止后立即有 (IRES)-cre 的定向插入。TH-Cre 大鼠可用于需要组织特异性表达的应用,包括光遗传学和转基因 floxed 系育种。

大鼠中的Sparsentan响应基因的翻译
保留所有权利。未经许可就不允许重复使用。(未经同行评审证明)是作者/资助者,他已授予Medrxiv的许可证,以永久显示预印本。此预印本版的版权持有人于2025年2月27日发布。 https://doi.org/10.1101/2025.02.26.25322958 doi:medrxiv preprint

昼夜节律的改变作为大鼠抑郁>的模型 div>
使用图像,视频片段和其他是版权保护主题的材料,将专门用于教育和信息性目的,必须引用他通过提及作者或作者获得的来源。 div>任何不同的用途,例如利润,复制,版本或修改,都将受到相应版权持有人的迫害和批准。 div>

大鼠神经母细胞瘤细胞系在无血清
合成培养基通常需要补充血清来支持培养细胞的增殖或存活。然而,血清的加入可能会显著影响实验的可重复性,因为即使是在混合血清样本中,由于供体的年龄、性别、营养和生理状态的差异,批次也会发生变化。此外,血清复杂而不确定的性质也使评估调节剂(如激素或神经递质)对培养神经系统细胞分化特性的影响变得复杂。这对于长期研究尤其重要,因为如果删除血清,细胞数量可能会大幅减少,在许多情况下,细胞可能会在数小时或数天内完全死亡。为了避免这些问题,已经对几种细胞系进行了改造,使其能够在无血清培养基中增殖(1-4)。然而,成功适应这些条件通常需要很长的时间,而且很少有细胞系能够表达分化功能。此外,这些适应性细胞系很可能已经丧失了一些更有趣的生理特性,例如对血清激素的反应。本实验室最近成功采用了另一种方法,即在无血清实验培养物中添加激素和生长因子。由于细胞从含血清培养基转移到无血清补充培养基后会继续生长,几乎没有或根本没有因转移而导致的滞后,因此似乎不涉及选择或适应。迄今为止,大鼠垂体 GH3、人宫颈 HeLa 细胞、小鼠黑色素瘤 M2R、小鼠胚胎癌 PCC.4 aza-1 和 F9、大鼠神经胶质瘤 C6、小鼠成纤维细胞 BALB 3T3 和 Swiss 3T3、正常大鼠滤泡 RF-1 以及正常睾丸 TM4 细胞系已被证明能在补充无血清培养基中增殖 (5-8)。 B104 大鼠神经母细胞瘤是一种中枢神经系统来源的细胞系,它表现出分化神经元的许多特性,例如动作电位的产生、神经递质的合成以及神经递质的存在

apelin-13对雄性大鼠的焦虑行为的影响
AIM:众所周知,Apelin-13是主要神经肽之一,在与情绪障碍有关的电路中起着明确而至关重要的作用。在病理焦虑的发展和/或维持中,海马和杏仁核的异常起着重要作用。在这里,我们评估了Apelin-13对雄性大鼠焦虑样行为的潜在抗焦虑作用。材料和方法:总共48名男性Wistar大鼠分为4组(n = 12)。控制(C),社会隔离(SI),Apelin-13(a)和社会隔离 + Apelin- 13(SI + A)。在C和A组中,每个笼子中有四只动物持续8周。 在Si和Si+A组中,将每只动物单独饲养8周。 在该APELIN-13给药之后,通过渗透泛滥。 使用高架迷宫(EPM),开放式测试(OFT)和灯光盒(LDB)评估了与焦虑/抑郁相关的行为。 我们还测量了APELIN-13,APELIN受体(APJ),脑衍生的神经营养因子(BDNF),哺乳动物阿托纳同源物1(Mash1),Nestin,Doublecortin(DCX)和神经素的表达。 这些是指示海马中焦虑机制的重要标记。 结果:我们的研究结果表明,Apelin-13给药减少了焦虑行为。 敞开的手臂完整,在A组中花费的时间更高。 在开放式测试中,SI组的修饰和饲养较低。 此外,在A组中,APELIN-13和APJ基因表达较高。在C和A组中,每个笼子中有四只动物持续8周。在Si和Si+A组中,将每只动物单独饲养8周。在该APELIN-13给药之后,通过渗透泛滥。使用高架迷宫(EPM),开放式测试(OFT)和灯光盒(LDB)评估了与焦虑/抑郁相关的行为。我们还测量了APELIN-13,APELIN受体(APJ),脑衍生的神经营养因子(BDNF),哺乳动物阿托纳同源物1(Mash1),Nestin,Doublecortin(DCX)和神经素的表达。这些是指示海马中焦虑机制的重要标记。结果:我们的研究结果表明,Apelin-13给药减少了焦虑行为。敞开的手臂完整,在A组中花费的时间更高。在开放式测试中,SI组的修饰和饲养较低。此外,在A组中,APELIN-13和APJ基因表达较高。结论:研究的结果表明,Apelin-13输注可能导致雄性大鼠与焦虑相关的行为减少。

FTY720在2型糖尿病大鼠中的影响
提高锂离子电池性能的关键是精确阐明电池的时间和空间层次结构。锂离子电池由阴极和阳极和含有电解质的分离器组成。锂离子电池的阴极和阳极由由活性材料,导电材料和粘合剂组成的复合材料制成,形成复杂的三维结构。由于锂离子反复插入并从活性材料中取出时,反应进行。因此,由于离子扩散而对活性材料的晶格进行了重组,从而导致相变。在活性材料 - 电解质界面上,锂离子的插入和去插入以及电荷转移反应进行。由于多种现象的相互作用,锂离子电池的电荷 - 放电反应是一个非平衡状态。在常规电池研究中执行的灾难性电池后的分析并不能准确理解反应速率和降解机制的主要因素。本综述介绍了有关锂离子电池的时间和空间层次结构的研究结果,重点是在电荷 - 解散反应期间进行的操作测量。第1章概述了锂离子电池的分层反应机理。第2章介绍了Operando测量技术,这对于分析很有用。第3章描述了电极 - 电解质界面的反应,即反应场,第4章讨论了由有源材料中的两相反应引起的非平衡结构变化。第5章介绍了复合电极的独特反应异质性的研究,该反应异质性可以实用。了解分层反应机制将为锂离子电池和下一代电池的设计提供有用的信息。
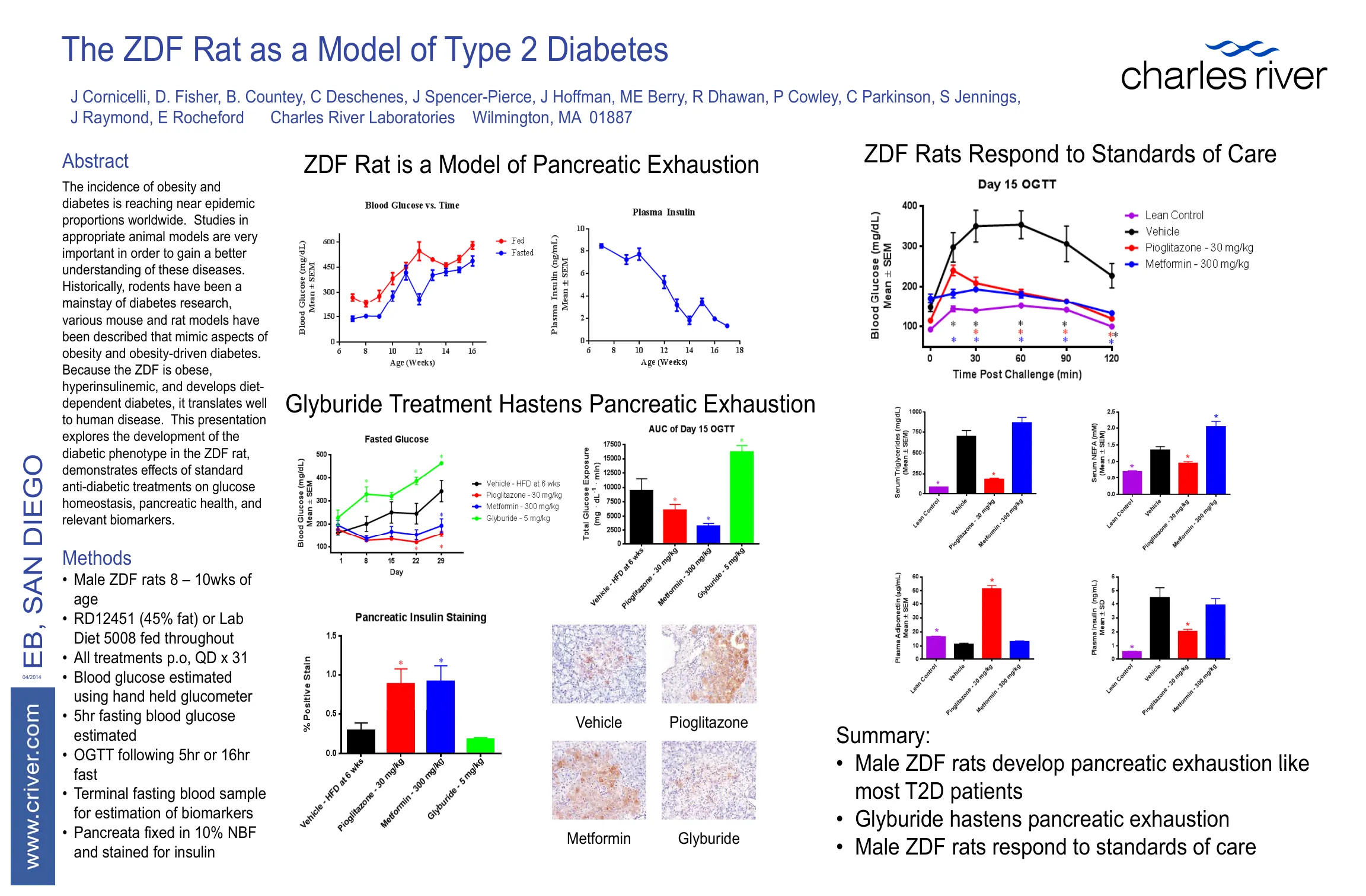
ZDF 大鼠作为 2 型糖尿病的模型 - NET
J Cornicelli、D. Fisher、B. Countey、C Deschenes、J Spencer-Pierce、J Hoffman、ME Berry、R Dhawan、P Cowley、C Parkinson、S Jennings、J Raymond、E Rocheford Charles River Laboratories 马萨诸塞州威尔明顿 01887

大鼠轻度爆炸性脑损伤后的社会心理损害
† 通讯作者 Riyi Shi,医学博士,哲学博士,625 Harrison St,West Lafayette,IN 47907,电话:(765)-494-6446,传真:(765)-494-7605,riyi@purdue.edu,William A. Truitt,哲学博士,320 W 15th St Office #314G,Indianapolis,IN 46202,电话:(317)-278-9050,btruitt@iu.edu。 * 同等贡献作者 作者贡献(CRediT 声明) Nicholas Race:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Katharine Andrews:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Elizabeth Lungwitz:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Sasha Vega-Alvarez:概念化、方法论、调查、验证、分析、写作 - 原始草稿和修订、可视化 Timothy Warner:调查、分析、写作 - 修订 Glen Acosta:调查、分析、写作 - 修订 Jiayue Cao:调查、分析、写作 - 修订 Kun-han Lu:调查、分析、写作 - 修订 Zhongming Liu:概念化、方法论、分析、写作 - 修订、监督 Amy Dietrich:调查、验证、分析、写作 - 修订 Sreeparna Majumdar:调查、验证、分析、写作-修改 Anantha Shekhar:分析、写作-修改、监督 William Truitt:概念化、方法论、分析、写作-修改、监督 Riyi Shi:概念化、方法论、分析、写作-修改、监督