XiaoMi-AI文件搜索系统
World File Search System天内结

老年女性 60 天内心力衰竭再入院率
背景:心力衰竭 (HF) 在患有心力衰竭保留射血分数 (HFpEF) 或心力衰竭降低射血分数 (HFrEF) 的老年人中普遍存在,并且早期 HF 再入院率很高。预防早期再入院很复杂,因为 HF 的两种亚型之间存在很大差异,并且预测模型不足以识别关键促成因素。目标:介绍有关选定临床、血流动力学、社会因素与 HFpEF 和 HFrEF 老年人早期(60 天)HF 再入院之间关系的研究,推导出再入院预测因素的概念模型,并了解文献在多大程度上解决了老年女性中的这些预测因素。方法:在四个计算机数据库中搜索与早期 HF 再入院和老年人 HF 指数住院后相关的临床、血流动力学和社会因素的研究。结果:最终审查纳入了 21 篇全文文章,并按主题组织。大多数研究集中于早期(30 天)HF 再入院,对 31 至 60 天期间的关注有限。确定了影响早期 HF 再入院的具体临床、血流动力学和社会因素。现有文献证实,影响指数 HF 住院后早期(60 天)HF 再入院的风险预测因子或其组合仍然不一致。此外,文献未能仅捕捉到这些预测因子对老年女性的影响。提出了一个风险预测因子的概念模型用于临床干预。结论:需要进一步评估以了解老年女性早期(31 至 60 天)HF 再入院的风险预测因子。© 2022 作者。由 Elsevier Inc. 出版。这是一篇根据 CC BY-NC-ND 许可协议开放获取的文章 ( http://creativecommons.org/licenses/by-nc-nd/4.0/ )


突变型结直肠癌
摘要 ◥ 目的:恩科拉非尼 + 西妥昔单抗 (E + C) 是治疗化疗难治性 BRAF V600E 转移性结直肠癌 (mCRC) 的有效选择。然而,有必要提高这种分子靶向治疗的疗效,并评估适合未治疗的 BRAF V600E mCRC 患者的方案。实验设计:我们使用 BRAF V600E mCRC 肿瘤异种移植进行了一系列体内研究。小鼠随机接受 5-氟尿嘧啶 (5-FU)、伊立替康或奥沙利铂方案 (FOLFIRI 或 FOLFOX)、(E + C) 或组合治疗。患者接受长期治疗直至病情进展,并使用降级策略模拟维持治疗。评估了细胞毒化疗或靶向治疗进展后的转录组变化。结果:FOLFIRI 或 E + C 作为一线治疗的抗肿瘤活性优于二线治疗,细胞毒性方案和靶向治疗之间存在部分交叉耐药性。

结直肠癌 - FDA
其他有效的筛查方法包括粪便检查(可配合或不配合柔性乙状结肠镜检查),柔性乙状结肠镜检查类似于结肠镜检查,但不能检查整个结肠。CT 结肠镜检查(放射科医生进行的检查)也用于筛查患者是否患有结肠直肠癌。如果发现异常,您的医生将确定是否需要进行后续结肠镜检查。

JT010N065SED/CED/FED/WED
* 连续集电极电流由最高结温限制。 *Collector current limited by maximum Junction temperature. 注 1 :脉冲宽度由最高结温限制。 Note1 : The pulse width is limited by the maximum junction temperature.



一天内构建多重阵列以利用天然 CRISPR 系统
摘要 CRISPR-Cas 系统是一种原核免疫系统,不仅在细菌和古细菌中广泛增殖,而且最近在人类生物学研究和应用中也广泛存在。迄今为止,许多工作都利用了合成的 sgRNA 和 CRISPR 核酸酶 Cas9,但阵列处理核酸酶的发现现在允许在异源宿主以及具有内源系统的生物体中使用更紧凑、天然的 CRISPR 阵列。不幸的是,多重天然 CRISPR 阵列的构建在技术上仍然具有挑战性、成本高昂和/或耗时。这种限制阻碍了在天然和异源宿主中涉及天然 CRISPR 阵列的研究。为了解决这个问题,我们提出了一种组装 CRISPR 阵列的方法,这种方法简单、快速、经济实惠且高度可扩展——我们用一天的工作组装了 9 间隔阵列。我们利用这种方法来利用高能力细菌 Acinetobacter baylyi 的内源性 CRISPR 系统,结果表明虽然单个间隔区并不总是能够完全有效地阻止通过天然能力获取 DNA,但多重天然 CRISPR 阵列可以实现几乎完全的 DNA 排除和基因组编辑,包括两者的多个靶标。除了展示一种将使各种应用受益的 CRISPR 阵列组装方法之外,我们还发现了一种潜在的对冲策略,用于平衡 CRISPR 防御与天然能力的 A. baylyi 中的 DNA 获取。CRISPR(成簇的规律间隔的短回文重复序列)-Cas 系统是一种适应性免疫机制,通常通过检测和切割确定的靶序列 1 来保护细菌和古菌免受核酸入侵。CRISPR 系统包括 Cas(CRISPR 相关)蛋白,以及与同样短的 DNA 间隔区交替的同名短直接重复序列阵列。间隔序列被转录成较长的前体crRNA,然后前体crRNA被加工成单个crRNA(CRISPR RNA),每个crRNA由与特定核酸靶标互补的单个间隔序列以及通常源自重复序列的发夹柄组成。这些crRNA与Cas效应蛋白(如Cas9)或蛋白质复合物(如CASCADE)结合。一旦结合,它们就会根据系统引导效应子到互补的DNA或RNA上,效应子通常会切割这些DNA或RNA。很快,许多实验室就将CRISPR介导的DNA切割应用于从精确的基因组工程到基因回路2到靶向的细菌菌株去除3-6等各种应用。自扩散的CRISPR构建体也被用于快速产生纯合二倍体敲除(诱变链式反应)7,初步研究表明

一天内构建多重阵列以利用天然 CRISPR-Cas 系统
摘要:CRISPR-Cas 系统是一种原核生物免疫系统,不仅在细菌和古菌中广泛存在,而且最近也在人类生物学研究和应用中得到应用。迄今为止,许多研究都利用了合成的 sgRNA 和 CRISPR 核酸酶 Cas9,但阵列处理核酸酶的发现现在允许在异源宿主以及具有内源系统的生物体中使用更紧凑、天然的 CRISPR 阵列。不幸的是,多重天然 CRISPR 阵列的构建在技术上仍然具有挑战性、成本高昂和/或耗时。这种限制阻碍了在天然和异源宿主中涉及天然 CRISPR 阵列的研究。为了解决这个问题,我们提出了一种组装 CRISPR 阵列的方法,该方法简单、快速、经济实惠且高度可扩展我们用 1 天的工作时间组装了 9 间隔阵列。我们利用这种方法来利用高能力细菌 Acinetobacter baylyi 的内源性 CRISPR-Cas 系统,结果表明,虽然单个间隔物并不总是能够完全有效地阻止通过天然能力获取 DNA,但多重天然 CRISPR 阵列可以实现几乎完全的 DNA 排除和基因组编辑,包括对两者的多个目标。除了展示一种将有益于各种应用的 CRISPR 阵列组装方法外,我们还发现了一种潜在的双赢策略,用于在天然能力的 A. baylyi 中平衡 CRISPR 防御与 DNA 获取。
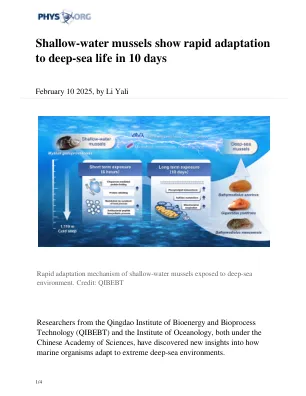
浅水贻贝在10天内快速适应深海寿命
值得注意的是,深海贻贝中的甲烷营养细菌 - 钥匙共生体 - 在暴露的浅水贻贝中占主导地位。这种转移与与免疫反应和内吞作用有关的基因表达的变化相关,突出了贻贝及其共生体之间的协同关系。