XiaoMi-AI文件搜索系统
World File Search System果蝇

光遗传学激活果蝇幼虫中巨型 GABA 能中枢脑中间神经元的奖励能力
1 莱布尼茨神经生物学研究所,学习和记忆遗传学系,马格德堡,39118,德国,2 莱比锡大学生物研究所动物生理学系,莱比锡,04103,德国,3 莱比锡大学生物研究所遗传学系,莱比锡,04103,德国,4 魏茨曼科学研究所分子细胞生物学系,雷霍沃特,7610001,以色列,5 亚琛工业大学成像和计算机视觉研究所,亚琛,52074,德国,6 波多黎各大学医学科学园区神经生物学研究所,旧圣胡安,波多黎各,00901,7 剑桥大学生理学、发育和神经科学系,剑桥,CB2 3EL,英国,8 珍妮莉亚研究园区,霍华德休斯医学研究所,阿什本, 20147,弗吉尼亚州,9 莱布尼茨神经生物学研究所,组合神经影像核心设施,马格德堡,39118,德国,10 加利福尼亚大学,分子,细胞和发育生物学系,加利福尼亚州洛杉矶 90095-1606,11 巴黎萨克雷大学,国立科学研究中心,巴黎萨克雷神经科学研究所,萨克雷,91400,法国,12 行为脑科学中心,马格德堡,39106,德国,13 奥托冯格里克大学生物学研究所,马格德堡,39120,德国

阿尔茨海默氏病和肠脑轴:果蝇杂物作为模型
研究表明,到2050年,由于淀粉样蛋白和tau蛋白的积累,有超过1.5亿人将患有阿尔茨海默氏病(AD)。除了遗传背景,内分泌干扰和细胞衰老外,肠道菌群的管理还成为AD诊断,进展和治疗的关键因素,因为某些细菌代谢物可以通过血液流过血流并跨越血脑屏障。这个迷你审查探讨了果蝇果蝇中的tau蛋白积累与肠道营养不良之间的关系。该模型有助于研究肠道衍生的代谢产物如何促进神经认知障碍和痴呆症。了解直接和间接细菌副产品(例如乳酸和乙酸酯)在神经胶质细胞活化和TAU蛋白动力学中的作用可能会提供对AD进展机制的见解,并有助于更有效的治疗方法。在这里,我们讨论了果蝇的简单性和广泛的遗传工具如何使其成为研究这些相互作用和测试潜在疗法(包括益生菌)的宝贵模型。将果蝇研究与其他已建立的模型整合在一起可能会揭示保守的途径,并加速发现为临床应用的翻译。

果蝇 yakuba 中 eIF4E1 直系同源物的基因模型
(A) 果蝇 (Drosophila melanogaster) 和果蝇 (D. yakuba) 中 eIF4E1 基因组邻域的同源性比较。细箭头表示果蝇 (D. melanogaster) (顶部) 和果蝇 (D. yakuba) (底部) 基因组中参考基因 eIF4E1 所在的 DNA 链。指向右侧的细箭头表示 eIF4E1 在果蝇 (D. melanogaster) 中位于正 (+) 链上,指向左侧的细箭头表示 eIF4E1 在果蝇 (D. yakuba) 中位于负 (-) 链上。指向与 eIF4E1 相同方向的宽基因箭头相对于细箭头位于同一链上,而指向与 eIF4E1 相反方向的宽基因箭头相对于细箭头位于相反链上。果蝇 (D. yakuba) 中的白色基因箭头表示与果蝇 (D. melanogaster) 中相应基因的直系同源。 D. yakuba 基因箭头中给出的基因符号表示 D. melanogaster 中的直系同源基因,而基因座标识符特定于 D. yakuba。(B)GEP UCSC Track Data Hub 中的基因模型(Raney 等人,2014 年)。D. yakuba 中 eIF4E1 的编码区显示在用户提供的 Track(黑色)中;CDS 用粗矩形表示,内含子用细线表示,箭头表示转录方向。后续证据轨迹包括 NCBI RefSeq 基因的 BLAT 比对(深蓝色,D. yakuba 的 Ref-Seq 基因比对)、D. melanogaster 蛋白质的 Spaln(紫色,D. melanogaster 的 Ref-Seq 蛋白质比对)、TransDecoder 预测的转录本和编码区(深绿色)、成年雌性和成年雄性的 RNA-Seq(分别为红色和浅蓝色;D. yakuba 的 Illumina RNA-Seq 读段比对)以及使用 D. yakuba RNA-Seq (SRP006203 - Graveley et al, 2010) 通过 regtools 预测的剪接点。显示的剪接点分别具有 232、500-999 和 >1000 的读取深度,支持读取为粉色、棕色和红色。 (C) 果蝇 (D. melanogaster) 中的 eIF4E1-PB (x 轴) 与果蝇 (D. yakuba) 中的直系同源肽 (y 轴) 的点图。左侧和底部表示氨基酸编号;顶部和右侧表示 CDS 编号,CDS 也以交替颜色突出显示。序列相似性降低的区域用红色圈出。 (D) 果蝇 (D. melanogaster) 中的 eIF4E1-PC (x 轴) 与果蝇 (D. yakuba) 中的直系同源肽 (y 轴) 的点图。序列相似性降低的区域用红色圈出。
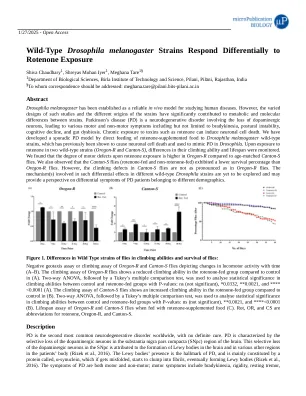
野生型果蝇melanogaster菌株对紫藤酮暴露的反应差异
果蝇Melanogaster已被确立为研究人类疾病的可靠模型。然而,此类研究的各种设计以及菌株的不同起源显着导致菌株之间的代谢和分子差异。帕金森氏病(PD)是一种神经退行性疾病,涉及多巴胺能神经元的丧失,导致各种运动和非运动症状,包括但不限于Bradykinesia,姿势不稳定,认知能力下降,认知能力下降和胆汁性营养不良。长期暴露于毒素(例如烤)可以诱导神经元细胞死亡。我们通过直接喂养烤烤面包酮的食物向果蝇Melanogaster野生型菌株开发了一种零星的PD模型,以前已证明该菌株会导致神经元细胞死亡,并用于模仿果蝇中的PD。在两种野生型菌株(俄勒冈-R和Canton-S)中暴露于鱼藤酮后,监测其攀爬能力和寿命的差异。我们发现,与年龄匹配的广州苍蝇相比,俄勒冈-R紫红酮暴露时的运动缺陷程度更高。我们还观察到,与俄勒冈-r蝇相比,广州蝇(烤面包酮喂养的和非洛诺酮)的生存百分比较低。但是,广州蝇中的攀爬缺陷并不像俄勒冈-r蝇中那样明显。在不同野生型果蝇菌株中涉及这种差异效应的机制尚待探索,并可能对属于不同人口统计学的PD患者的差异症状提供视角。

主要物种决定果蝇 - 寄生物食物网络动力学和稳定性比复杂性,变暖或入侵更强烈
。cc-by-nc-nd 4.0国际许可证。是根据作者/资助者提供的预印本(未经同行评审认证)提供的,他已授予Biorxiv的许可证,以在2025年1月27日发布的此版本中显示此版本的版权持有人。 https://doi.org/10.1101/2025.01.27.634676 doi:Biorxiv Preprint

自噬依赖的丝状伪足动力学限制了果蝇大脑连接过程中突触伙伴的选择
大脑连接非常精确,但大多数神经元一旦有机会就会与错误的伙伴形成突触。动态轴突-树突定位可以限制突触形成相遇,但发育中的大脑中时空相互作用动力学及其调节仍然基本未知。在这里,我们表明轴突伪足的动力学限制了突触形成和伙伴选择,而这些神经元原本不会被阻止形成错误的突触。利用 4D 成像技术对发育中的果蝇大脑进行研究,我们发现伪足动力学受自噬调控,自噬是一种普遍的降解机制,其在大脑发育中的作用仍不太清楚。自噬体以令人惊讶的特殊性在突触形成伪足中形成,随后伪足崩塌。计算建模和遗传实验表明,突触构建材料的自噬降解改变定量调节突触形成。伪足稳定性的增加导致错误的突触伙伴关系。因此,自噬通过动力学排除过程来限制不适当的伴侣选择,这对于连接特异性至关重要。

细胞周期依赖性提示调节果蝇中央脑神经干细胞的时间模式
在神经系统发育过程中,不同类型的神经元和神经胶质是由自我更新神经干细胞(NSC)依次产生的。NSC中基因表达的时间变化被认为调节神经di versity。但是,调节这些时间基因过渡的时机的机制仍然很少理解。果蝇II型NSC,例如人类外部radial胶质神经胶质,分裂为自我更新并产生中间神经祖细胞,扩大和多样化神经元的群体,该神经元的种群神经支配了中央复合体,这是一种脑部的大脑区域。II型NSC在暂时的十几个基因上表达,广泛地分类为早期和晚期基因。一个保守的基因,通过激活ecdysone受体(ECR)表达,七个UP介导了早期至晚期的压缩。然而,决定了ECR表达的时间,因此,尚不清楚基因转变。这项研究提出了细胞周期进程和细胞因子的固有机制是否需要诱导NSC早期脑结构过渡。通过加入释放NSC细胞周期或阻断细胞因子的突变克隆,我们表明这两个过程对于早期到偏移过渡都是必需的。当NSC是细胞周期或抑制了Cyto kinesis时,早期的基因IMP未能下调并持续到旧的NSC中,而晚期因素ECR和Syncrip未能表达出来。此外,我们表明,早期的七个因素不足以

研究文章捕获了果蝇的连续,长时间的行为变化
动物行为跨越了许多时间尺度,从短时,秒的动作到每天的节奏,到衰老期间的终生变化。为了访问较长的行为时间尺度,我们以每秒100帧的速度记录了单个果蝇Melanogaster,每次在蔗糖媒体上的无特色竞技场上一次最多7天。我们使用深度学习框架切片为47个人生产全身姿势数据集,导致近20亿个姿势实例。我们确定了陈规定型的行为,例如修饰,长鼻延伸和运动,并使用所得的伦理图来探索苍蝇的行为在实验中的一天和几天之间的变化。我们在所有定型行为中发现不同的每日模式,添加了有关不同修饰方式,长鼻延长持续时间和运动速度的趋势的特定信息。Melanogaster昼夜节律周期。使用我们对行为的整体测量,我们发现黎明后的小时是苍蝇日常行为模式中的独特时间点,并且这个小时的行为组成与其他健康指标(例如运动速度和时间的一部分时间花费移动与休息)都很好地跟踪。此处介绍的方法,数据和分析为我们提供了d的新图片。Melanogaster跨时标的行为,揭示了暗示未探索潜在生物学机制的新型特征。

果蝇中转录起始位点的注释
引入尽管居住在具有高重复密度的域中,但果蝇Melanogaster muller f元素基因还是在与正念基因相同的定量范围内表达(Riddle等人。2012)。比较Muller F和D元素基因的转录起始位点(TSS)附近的基序的类型和分布可以帮助阐明使Muller F元素基因在异性域中起作用的因素。主题分析的第一步是产生TSS的高质量注释,以定义搜索保守基序的区域。TSS的比较注释比编码区域的注释更具挑战性,因为5'和3'未翻译区域(UTR)的发展比编码区域更快,并且提供了支持注释的外部证据较少。例如,大多数基因查找器仅预测编码区域,而RNA-seq读取覆盖率数据通常没有提供足够的证据来推断TSS的精确位置。因此,与编码区域的注释相比,TSS的注释具有更高的不确定性程度。在某些情况下,我们可能只能定义一个可以找到TSS的基因组区域。本演练将说明使用D. biarmipes muller f element Project contig35 [8月。 2013(GEP/DOT)组件]。

果蝇表皮细胞本质上是机械敏感的,调节伤害性行为输出
。cc-by 4.0国际许可(未经Peer Review尚未获得认证)是作者/资助者,他已授予Biorxiv的许可证,以永久显示预印本。这是该版本的版权持有人,该版本发布于2025年1月7日。 https://doi.org/10.1101/2022.10.07.511265 doi:Biorxiv Preprint