XiaoMi-AI文件搜索系统
World File Search System标的
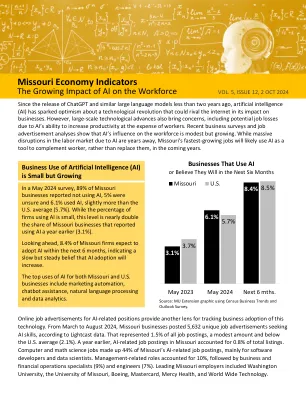
人工智能对密苏里州经济指标的影响日益增大
人工智能相关职位的在线招聘广告为追踪企业采用这项技术提供了另一个视角。根据 Lightcast 的数据,从 2024 年 3 月到 8 月,密苏里州的企业发布了 5,632 条寻求人工智能技能的独特招聘广告。这占所有招聘信息的 1.5%,数量不多,低于美国平均水平(2.1%)。一年前,密苏里州的人工智能相关职位占总招聘信息的 0.8%。计算机和数学科学职位占密苏里州人工智能相关职位的 44%,主要针对软件开发人员和数据科学家。管理相关职位占 10%,其次是商业和金融运营专家(9%)和工程师(7%)。密苏里州的主要雇主包括华盛顿大学、密苏里大学、波音公司、万事达卡、Mercy Health 和 World Wide Technology。

数据驱动的表征临床,神经影像学和基于血液的指标的创伤性脑损伤严重程度
使用格拉斯哥昏迷量表(GCS)总分具有众所周知的局限性,将创伤性脑损伤(TBI)表征为轻度,中度或重度的常规临床方法,促使人们呼吁采用更复杂的策略来表征TBI。在这里,我们使用项目响应理论(IRT)来开发一种新的方法来量化TBI严重程度,该方法结合了神经影像学和基于血液的生物标志物以及临床指标。在TBI(TRACK-TBI)研究样本(n = 2545)的多中心转化研究和临床知识中,我们表明,一组23套临床,头部计算机断层扫描(CT)和基于血液的生物标志物变量熟悉临床医生和研究人员熟悉的生物标志物变量。我们说明了如何使用IRT来识别这些特征的相对值,以估计个人沿TBI严重性连续体的位置。最后,我们表明,使用这种基于IRT的新方法产生的TBI严重程度得分逐渐预测了经典临床(轻度,中,严重或国际任务)预后和分析TBI(影响)分类方法的临床试验的功能结果。我们的调查结果直接为正在进行的国际努力提供了完善和部署新的务实的,经验上支持TBI的策略,同时说明了一种可能有助于发展其他疾病的分期系统有用的策略。

OMS 观点和 OMS 国家内 RA 指标的使用
MISO 关税模块 E-1 规定:“本模块 E-1 中的任何内容均不影响各州对额外容量建设的现有管辖权或各州制定和执行充足性标准的权力。本模块 E-1 中的资源充足性要求 (RAR) 无意且不会以任何方式影响各州对其管辖范围内实体的行动。”

巨噬细胞可塑性对炎症性关节炎的贡献及其作为治疗靶标的潜力
摘要:炎症性关节炎是常见的慢性炎症自身免疫性疾病,这些疾病因进行性,破坏性的炎症而导致功能丧失和显着合并症的关节疾病。重要的是,没有治疗方法,只有20%的患者在2年以上实现无药缓解。巨噬细胞在维持体内平衡方面起着至关重要的作用,但是,在错误的环境线索下,巨噬细胞成为慢性滑膜炎症的驱动因素。基于当前的“教条”,M1巨噬细胞分泌促炎性细胞因子和趋化因子,促进组织降解,关节和骨侵蚀,这会导致疾病进展加速。另一方面,M2巨噬细胞分泌与伤口愈合,组织重塑和炎症分辨率相关的抗炎介质。目前,已经鉴定出四种亚类型M2巨噬细胞,即M2A,M2B,M2C和M2D。然而,由于巨噬细胞的可塑性和重极化的能力,可能存在更多的亚型。巨噬细胞是高度塑性的,极化作为具有不同中间表型的连续体存在。这种可塑性是通过响应环境刺激和新陈代谢转移的高度正态性基因组来实现的。在疾病早期阶段开始治疗对于证明的预后和患者预后很重要。目前,没有专门针对巨噬细胞的治疗方法。正在进行的临床试验中正在研究此类治疗剂。已经提出,促炎性巨噬细胞对抗炎表型的复制是作为靶向M1/M2不平衡的有效方法,反过来又是IA疾病的潜在治疗策略。因此,阐明控制巨噬细胞可塑性的机制对于新型巨噬细胞靶向治疗剂的成功至关重要。

英国政府的目标是到 2035 年利用太阳能发电 70 吉瓦。迅速实现这一目标将是实现目标的重要组成部分
1 根据规划,我们假设这三个太阳能发电场将占用 35 公顷 2 级农业用地、292 公顷 3a 级土地、1,339 公顷 3b 级土地和 411 公顷 4 级土地(参见:Gate Burton、Mallard Pass 和 Sunnica)。这些计划表明,农业生产力最高的 1 级土地不受影响。我们假设 2 级和 3a 级土地是可耕地(英格兰的可耕地面积大约等于英格兰 1、2 和 3a 级土地的面积),动物在 3b 级和 4 级土地上放牧。因此,我们根据国家粮食战略的分析估算了这些地区的产量损失百分比,即英国 84% 的卡路里产量来自 37% 的可耕地,16% 的卡路里产量来自 63% 的放牧农田。根据英国每人每天的卡路里供应量估计为 3.344 千卡,其中 49% 的卡路里是在国内生产的,我们计算出这三个太阳能发电场将损失多少卡路里。相比之下,使用标准的能量密度估计值,2024 年小麦产量与 2023 年相比减少了 27%,这比三个太阳能发电场造成的粮食损失高出 5,761 倍。
![b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A](/simg/f/f6a2bb5f504472f4ca10c1ecebfe7702878cdb55.webp)
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。Song,线粒体DNA:分布,突变和消除,细胞,8(2019)。 [3] F. Liu,D.E。 Sanin,X。Wang,肺癌中的线粒体DNA,实验医学和生物学进展,1038(2017)9-22。 [4] J. Zhang,J。 Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 Jia,W.G。Li,D.S。Pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶标的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ XCAD \ Xada,M。Robledo,Primpol:DNA复制酶之间的突破性和潜在的癌症治疗靶标的癌症治疗,生物分子,12(20222)。 [7] A.O. Giacomelli,X。Yang,R.E。 Lintner,J.M。McFarland,M。Duby,J.Kim,T.P。 D.Y. Howard武田Ly,E。Kim,H.S。甘农(B. J.G. Aguirre Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。约翰内森,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。Gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。 [9] C.Y. Dai,C.C。 Ng,G.C.C。 Hung,I。Kirmes,L.A
b'[2] C. Yan,X。Duanmu,L。Zeng,B。Liu,Z。歌曲,线粒体DNA:分布,突变和消除,细胞,8(2019)。[3] F. Liu,D.E。Sanin,X。Wang,肺癌中的线粒体DNA,实验医学与生物学进展,1038(2017)9-22。[4] J. Zhang,J。[5] P.P.Jia,M。Junaid,Y.B。 MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Jia,M。Junaid,Y.B。MA,F。Ahmad,Y.F。 jia,W.G。 li,D.S。 pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。 [6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。 Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。 [7] A.O. Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。MA,F。Ahmad,Y.F。jia,W.G。li,D.S。pei,人类DNA2(HDNA2)作为癌症和其他疾病的潜在靶点的作用:系统评价,DNA修复(AMST),59(2017)9-19。[6] A. D \ XC3 \ Xadaz-Talavera,C。Montero-Conde,L.J。Leandro-Garc \ XC3 \ Xada,M。Robledo,Primpol:DNA复制酶的突破和潜在的癌症治疗新靶标,生物分子,12(2022)。[7] A.O.Giacomelli,X。Yang,R.E。 lintner,J.M. McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Giacomelli,X。Yang,R.E。lintner,J.M.McFarland,M。Duby,J。Kim,T.P。 D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。McFarland,M。Duby,J。Kim,T.P。D.Y. Howard Takeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。D.Y. HowardTakeda,S.H。 ly,E。Kim,H.S。 Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Takeda,S.H。ly,E。Kim,H.S。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。 Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Gannon,B。Hurhula,T。Sharpe,A。Goodale,B。Fritchman,S。Seelman,F。Vazquez,A。Tsherniak,A.J。Aguirre,J.G。 Doench,F。Piccioni,C.W.M。 Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Aguirre,J.G。Doench,F。Piccioni,C.W.M。Roberts,M。Meyerson,G。Getz,C.M。 Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Roberts,M。Meyerson,G。Getz,C.M。Johannessen,D.E。 根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。 [8] G.A. Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Johannessen,D.E。根,W.C。 Hahn,突变过程塑造了人类癌症中TP53突变的景观,NAT Genet,50(2018)1381-1387。[8] G.A.Fontana,H.L。 [9] C.Y. dai,C.C。 ng,G.C.C。 Hung,I。Kirmes,L.A。Hughes,Y。Fontana,H.L。[9] C.Y.dai,C.C。ng,G.C.C。Hung,I。Kirmes,L.A。Hughes,Y。gahlon,线粒体DNA缺失形成的复制和修复机制,核酸res,48(2020)11244-11258。du,C.A。Brosnan,A。Ahier,A。Hahn,C.M。 Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。 [10] L. Ou,H。Liu,C。Peng,Y. [11] H. Liu,J。Weng,C.L.H。 Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Brosnan,A。Ahier,A。Hahn,C.M。Haynes,O。Rackham,A。Filipovska,S。Zuryn,ATFS-1,通过促进转录修复,自然细胞生物学,25(2023)1111-1120来抵消线粒体DNA损伤。[10] L. Ou,H。Liu,C。Peng,Y.[11] H. Liu,J。Weng,C.L.H。Huang,A.P。 杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。 [12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。 [13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。。 [14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。 [15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。 [16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。 2025.529997。 [17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。Huang,A.P。杰克逊,癌症的电压门控钠通道,生物标志物研究,12(2024)70。[12] H. Liu,A。Dong,A.M。 Rasteh,P。Wang,J。Weng,乳腺癌中新型T细胞CD8 +标记的鉴定,Scientific Reports,14(2024)19142。[13] H. Liu,T。Tang,基于MAPK信号途径的胶质瘤亚型,机器学习风险模型和关键集线器蛋白识别,科学报告,13(2023)19055。[14] H. Liu,T。Tang,《泛滥成灾与基因集的泛癌遗传分析》,癌症遗传学,278-279(2023)91-103。[15] H. Liu,T。Tang,《胶质瘤IGFBP的生物信息学研究》,涉及其诊断,预后和治疗预测值,AM J Transl Res,15(2023)2140-2155。[16] H. Liu,T。Tang,《泛滥成灾基因套件的泛 - 癌遗传分析》,Biorxiv,(2023),2023.2002。2025.529997。[17] H. Liu,库糖凋亡在肾脏肾透明细胞癌中的表达和潜在免疫受累,癌症遗传学,274-275(2023)21-25。[18] L. Hengrui,《中药用于癌症治疗中使用的有毒药物的例子》,J Tradit Chin Med,43(2023)209-210。[19] H. Liu,J。Weng,《 Rad51的Pan-Cancer生物信息学分析》,涉及诊断,预后和治疗预测的值,肿瘤学的前沿,12(2022)。[20] H. Liu,J。Weng,胶质瘤中细胞周期蛋白依赖性激酶2(CDK2)的全面生物信息学分析,Gene,(2022)146325。[21] H. Liu,T。Tang,Pan-Cancer的库糖胞化和铜代谢相关的基因集,肿瘤学的边界,12(2022)952290。[22] H. Liu,Y。Li,Cornichon家族AMPA受体辅助蛋白4(CNIH4)在头部和颈部鳞状细胞癌中的潜在作用,癌症生物标志物:疾病标志物A部分(2022)。[23] H. Liu,J.P。Dilger,J。Lin,pan-Cancer-Biodorminicals-Informinical-Informicals Trpm7的文献综述,Pharmacol Ther(2022)108302。[24] H. Liu,cuproptosis Gene Set的Pan-Canter概况,《美国癌症研究杂志》,第12期(2022)4074-4081。[25] Y. Liu,H。Liu,氨基酰基TRNA合成酶复合物的临床能力相互作用多功能蛋白1(AIMP1),用于头颈鳞状细胞癌,癌症生物标志物:疾病标志物A节A节(20222)。[26] Y. Li,H。Liu,Y。Han,在头部和颈部鳞状细胞癌中,Cornichon家族AMPA受体辅助蛋白4(CNIH4)的潜在作用,研究方形(2021)。 '

全球目标的AI Gen AI
注意:1)2019-2022 =基于实际的复合年增长率。2023-2026 =自我报告的预期应力测试与分析师的期望。2)平均利润率(EBITDA/收入)基于2019 - 2022年的实际产品。我们进一步使用面板数据模型来测试重新发明年数之间的关系(从受访者报告采用其重新发明策略的一年开始)和EBITDA利润率(控制行业,地理和公司规模)。金融服务公司被排除在此分析之外。3)2021基于TER 1.0研究。2023-2026 =自我报告的期望。可持续性=来自Capital IQ的ESG数据。体验=客户关系管理的度量和S&P全球和事实供应链中的供应商流失。人才和I&d = net net更好的指标,ESG书籍和S&P Global。创新=有形和无形支出的效率。资料来源:埃森哲重新发明调查,2022年11月(n = 1,516)和10月至nov 2023(n = 1,500)

图标的能源效率和碳足迹 -
•大气动力学,对流为5个示踪剂•辐射(“ ecrad”)•云覆盖•湍流•饱和调节•graupel Microphysics•大气从初始状态旋转•1天模拟;各种分辨率,合奏填充1个机柜

对遗传学和基因组学机器学习的模型评估指标的综述
机器学习(ML)在遗传学和基因组学方面表现出了巨大的希望,在遗传学和基因组学中,大而复杂的数据集有可能洞悉疾病风险的许多方面,遗传疾病的发病机理以及对健康和健康的预测。但是,由于这种可能性,有责任谨慎行事,以防偏见和可能产生有害意外影响的结果的侵蚀。因此,研究人员必须了解用于评估ML模型的指标,这些指标可能影响结果的关键解释。在这篇评论中,我们提供了用于聚类,分类和回归的ML指标概述,并突出了每个指标的优点和缺点。我们还详细介绍了模型评估期间发生的常见陷阱。最后,我们提供了研究人员如何从基因组学角度进行评估和利用ML模型结果的示例。

更新移植后移植物存活率指标的标准
紧急工作组已设定了一个宏伟目标,即在 2026 年完成 60,000 例移植手术。为支持实现这一宏伟目标,工作组评估了可能对利用率和效率造成障碍的 OPTN 政策和章程。MPSC 对移植后移植物存活率的审查被确定为一个潜在障碍,因为人们认为 MPSC 对移植项目进行移植后移植物存活率审查的可能性会导致规避风险的行为。MPSC 不支持暂停对所有移植后结果的审查,理由是担心患者安全以及患者和公众的看法,并认识到 MPSC 对项目的审查可以推动积极的变化,从而增加对项目的机构支持。讨论了潜在的替代方案,包括考虑更改审查的标记阈值、风险调整模型改进和额外的教育工作。1 OPTN 执行委员会要求 MPSC 考虑调整移植后结果指标标记阈值,从而通过标记更少的项目来消除增加移植的障碍。 OPTN 执行委员会要求 MPSC 还考虑调整要约接受标记阈值,以涵盖更多项目,从而为移植项目接受更多器官创造更强的激励。MPSC 建议将成人移植接受者 90 天移植物存活率和 1 年(以 90 天移植物存活率为条件)的标记阈值从移植项目风险比大于 1.75(比预期高 75%)的 50% 概率更改为移植项目风险比大于 2.25(比预期高 125%)的 50% 概率。对于移植后移植物存活率指标,1.0 风险比表示移植项目在考虑到捐赠者和接受者特征的情况下表现符合预期,而更高的风险比表示项目表现比预期差。MPSC 指出,2.25 风险比的阈值将继续识别最需要改进的移植项目,并且大多数严重的患者安全问题都是通过其他监测活动发现的。这一阈值变化应能减少移植项目对 MPSC 潜在绩效监控的担忧,并鼓励移植项目接受和移植更复杂的供体器官,从而让更多候选人更快地接受移植。MPSC 决定不改变录取率标记阈值,因为自 2023 年 7 月实施该指标以来,还没有足够的时间来评估移植项目对录取率的审查效果,也为进一步开发适用于所有器官的强大多标准录取过滤器提供了机会。