XiaoMi-AI文件搜索系统
World File Search System着丝粒

两种隐龟的从头基因组组装
了解物种间染色质构象的进化对于阐明基因组的结构和可塑性至关重要。线性远距离基因座的非随机相互作用以物种特异性模式调节基因功能,影响基因组功能、进化,并最终影响物种形成。然而,来自非模式生物的数据很少。为了捕捉脊椎动物染色质构象的宏观进化多样性,我们通过 Illumina 测序、染色体构象捕获和 RNA 测序为两种隐颈龟 (cryptodiran,藏颈龟) 生成从头基因组组装:Apalone spinifera (ZZ/ZW,2 n = 66) 和 Staurotypus triporcatus (XX/XY,2 n = 54)。除了在线性基因组中检测到的融合/裂变事件外,我们还检测到龟类的三维 (3D) 染色质结构与其他羊膜动物存在差异。也就是说,全基因组比较揭示了龟类染色体重排的不同趋势:(1)鳖科(Trionychidae)的基因组改组率较低,而鸡(可能是龟类的祖先)与核型高度保守;(2)动胸龟科(Kinosternidae)和翠龟科(Emydidae)的融合/裂变率中等。此外,我们还发现了一种染色体折叠模式,这种模式使以前在龟类中未检测到的“着丝粒 - 端粒相互作用”成为可能。“着丝粒 - 端粒相互作用”(本文发现)加上“着丝粒聚集”(之前在蜥蜴类中报道过)的组合龟类模式对于羊膜动物来说是新颖的,它反驳了以前关于羊膜动物 3D 染色质结构的假设。我们假设,在龟类中发现的不同模式起源于羊膜动物祖先状态,该状态由核结构定义,微染色体之间存在广泛的关联,这些关联在线性基因组改组后得以保留。

植物染色体工程——过去、现在和未来
自发染色体重排 (CR) 在物种形成、基因组进化和作物驯化中起着至关重要的作用。为了能够利用 CR 的育种潜力,人们开始通过 X 射线照射将染色体片段化,从而进行植物染色体工程。随着 CRISPR/Cas 系统的兴起,人们可以高效地在任意染色体位置诱导双链断裂 (DSB)。这使得预先设计的染色体工程达到了全新的水平。可以通过诱导染色体易位来打破特定基因之间的遗传连锁。可以恢复抑制遗传交换的自然倒位以进行育种。此外,人们已经开发出各种通过缩小常规标准 A 染色体或额外 B 染色体来构建微型染色体的方法,这些方法可以作为未来植物生物技术的载体。最近,人们可以构建一个功能性的合成着丝粒。此外,人们已经建立了不同的基因组单倍体化方法,其中一些方法基于着丝粒操作。未来,我们期望看到更复杂的重组,这些重组可以与重组酶等先前开发的工程技术相结合。染色体工程可能有助于重新定义遗传连锁群、改变染色体数量、在微型载货染色体上堆叠有益基因,或建立遗传隔离以避免杂交。
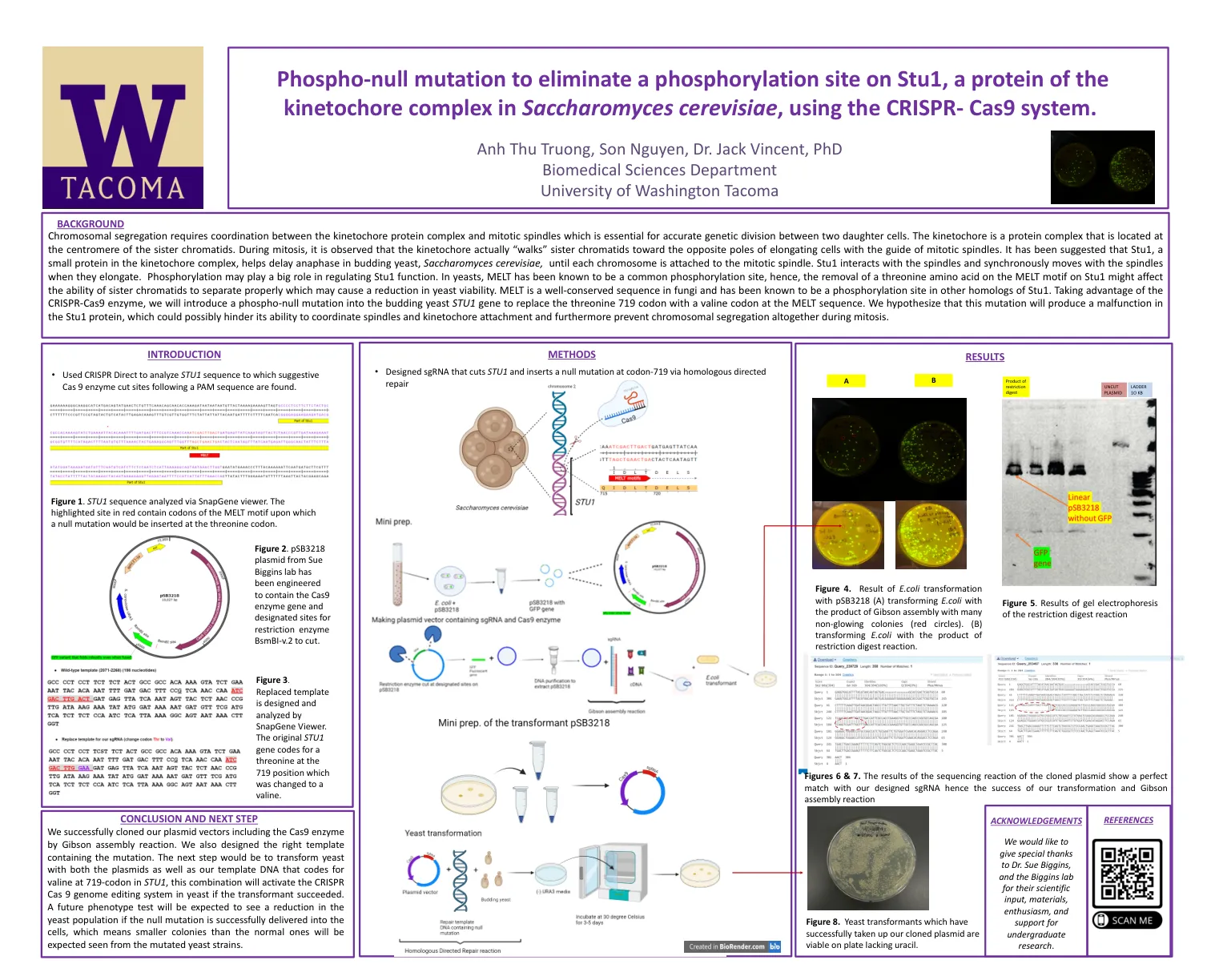
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

绘制完全合成植物染色体的路径
合成生物学的概念有可能改变植物遗传学,无论是在我们分析遗传途径的方式上,还是在我们将这些知识转化为有用应用的方式上。虽然合成生物学可以应用于单个基因或小群基因的水平,但本评论重点关注设计完全合成的植物染色体的最终挑战。这种规模的工程将使我们能够操纵整个基因组结构并同时修改多种途径和性状。基因组合成的进展使得植物染色体构建的初始阶段很可能发生在细菌和酵母中。在这里,我将讨论接下来的步骤,包括克服与植物转化、功能性着丝粒设计和确保准确的减数分裂传递相关的技术障碍的具体方法。

将细胞融合与 CRISPR-Cas9 编辑相结合,在酵母中克隆较大的 DNA 片段或完整的细菌基因组
图 1:CReasPy-Fusion 方法的实验流程示意图。步骤 1(借用 CReasPy-Cloning 策略,左栏):用两个质粒转化酵母,从而表达 Cas9 核酸酶和 gRNA。步骤 2(借用 Fusion Cloning 策略,右栏):在线性重组模板(由酵母元件 CEN-HIS3 组成,带有或不带有 ARS,两侧是与目标基因座两侧相同的两个重组臂和一个抗生素抗性标记)存在下,将预装 pCas9 和 pgRNA 的酵母细胞与支原体细胞接触。步骤 3:进入酵母细胞后,目标基因组被 Cas9 切割,随后由酵母同源重组系统使用提供的线性 DNA 片段作为模板进行修复。因此,细菌基因组现在包括插入到精确位置的酵母元素,并由酵母作为着丝粒质粒携带。

将细胞融合与 CRISPR-Cas9 编辑相结合,在酵母中克隆较大的 DNA 片段或完整的细菌基因组
图 1:CReasPy-Fusion 方法的实验流程示意图。步骤 1(借用 CReasPy-Cloning 策略,左栏):用两个质粒转化酵母,从而表达 Cas9 核酸酶和 gRNA。步骤 2(借用 Fusion Cloning 策略,右栏):在线性重组模板(由酵母元件 CEN-HIS3 组成,带有或不带有 ARS,两侧是与目标基因座两侧相同的两个重组臂和一个抗生素抗性标记)存在下,将预装 pCas9 和 pgRNA 的酵母细胞与支原体细胞接触。步骤 3:进入酵母细胞后,目标基因组被 Cas9 切割,随后由酵母同源重组系统使用提供的线性 DNA 片段作为模板进行修复。因此,细菌基因组现在包括插入到精确位置的酵母元素,并由酵母作为着丝粒质粒携带。

染色体外DNA(ecDNA):肿瘤异质性、基因组重塑和耐药性的起源。
癌细胞基因组含有正常细胞中没有的环状染色体外 DNA (ecDNA) 元素。临床样本分析表明,它们在大多数癌症中很常见,它们的存在预示着不良预后。它们通常含有高表达的增强子和驱动致癌基因。环状 ecDNA 拓扑结构导致染色质开放构象并产生新的基因调控相互作用,包括与远端增强子的相互作用。着丝粒的缺失导致细胞分裂过程中 ecDNA 随机分布,并且编码在其上的基因以非孟德尔方式传播。ecDNA 可以整合到染色体 DNA 中和退出。特定 ecDNA 的数量会随着治疗而改变。这种重塑癌症基因组的动态能力挑战了长期存在的基本原理,为肿瘤异质性、癌症基因组重塑和耐药性提供了新的见解。

深度强化学习的神经生物学
在哪里可以找到更多信息? Akera, T.、Trimm, E. 和 Lampson, MA (2019)。自私着丝粒减数分裂作弊的分子策略。Cell 178,1132–1144.e10。Burt, A. 和 Crisanti, A. (2018) 基因驱动:进化与合成。ACS Chem. Biol. 13,343–346。Cazemajor, M.、Joly, D. 和 Montchamp-Moreau, C. (2000)。拟果蝇的性别比例减数分裂驱动与 Y 染色体的方程不分离有关。Genetics 154,229–236。Crow, JF (1991)。孟德尔分离为何如此精确?BioEssays 13,305–312。 Dawe, RK, Lowry, EG, Gent, JI, Stitzer, MC, Swentowsky, KW, Higgins, DM, Ross-Ibarra, J., Wallace, JG, Kanizay, LB, Alabady, M., et al . (2018). 驱动蛋白-14 马达激活新着丝粒以促进玉米减数分裂驱动。Cell 173 , 839–850。Dyer, KA, Charlesworth, B., 和 Jaenike, J. (2007). 减数分裂驱动导致的染色体范围连锁不平衡。Proc. Natl. Acad. Sci. USA 104 , 1587–1592。Herrmann, BG, Koschorz, B., Wertz, K., McLaughlin, KJ, 和 Kispert, A. (1999)。 t 复合体反应基因编码的蛋白激酶导致非孟德尔遗传。自然 402,141–146。Larracuente, AM 和 Presgraves, DC (2012)。果蝇的自私分离扭曲基因复合体。遗传学 192,33–53。Lindholm, AK、Dyer, KA、Firman, RC、Fishman, L.、Forstmeier, W.、Holman, L.、Johannesson, H.、Knief, U.、Kokko, H.、Larracuente, AM 等人 (2016)。减数分裂驱动的生态学和进化动力学。生态学发展趋势 31,315–326。Sandler, L. 和 Novitski, E. (1957)。减数分裂驱动作为一种进化力量。美国自然。 91 , 105–110。Zanders, SE 和 Unckless, RL (2019)。减数分裂驱动因素的生育成本。Curr. Biol. 29 , R512– R520。

HJURP 作为癌症治疗靶点的新兴作用
霍利迪连接识别蛋白 (HJURP) 是着丝粒蛋白 A (CENP-A) 的关键分子伴侣,对有丝分裂期间的染色体分离和细胞周期调控至关重要。最近的研究已经确定了 HJURP 在致癌作用中的重要作用。在各种人类癌症中都观察到 HJURP 表达的异常上调,例如非小细胞肺癌 (NSCLC)、肝细胞癌 (HCC)、膀胱癌和乳腺癌,并且与不良的病理发展和预后有关。体外和体内研究表明,HJURP 主要通过调节细胞周期、细胞衰老和上皮-间质转化 (EMT) 发挥致癌功能。本综述旨在评估 HJURP 在人类癌症中的预后意义并总结针对 HJURP 的抗肿瘤研究。还讨论了调节 HJURP 在致癌作用中的因素及其相应的影响,以提供新的见解,以针对 HJURP 作为一种有前途的癌症治疗策略。

runx1家族血小板疾病概况人类基因组项目以外的基因组学和人类遗传学的年度综述:完整的人类基因组序列和Pangenome参考的时代
人类基因组项目是一个巨大的成就,为人类物种的遗传学和基因组学探索了无数的基础。多年来,人类基因组参考序列仍然不完整,并且缺乏人类遗传多样性的代表。最近,已经出现了两个重大进展来解决这些缺点:完全无间隙的人类基因组序列,例如由端粒到telomere群结的结合所开发的,以及高质量的pangenomes,例如由人类Pangenome Pangenome参考联盟中的dna序列组成和基因组合的依赖性,例如,由人类Pangenome PangeNome参考核心组成的核心和基因组合的核心,历史上难以顺序的区域,包括着丝粒,端粒和分段重复。同时,Pangenomes捕获了全世界种群中广泛的遗传多样性。共同发展了基因组学研究的新时代,增强了基因组分析的准确性,铺平了精确医学的道路,并有助于更深入地了解人类生物学。