XiaoMi-AI文件搜索系统
World File Search System种群

用高线粒体含量过滤细胞在癌症单细胞研究中去除可行的代谢改变的恶性细胞种群
此外,PCTMT已与细胞特异性代谢活性密切相关,从而导致不同细胞类型的实质性变异性,并且经常超过传统过滤器设定的阈值[8,14,16,17]。例如,蒙特塞拉特 - 尤索索(Montserrat-Ayuso)和埃斯特维(Esteve-Codina)[12]认为,常规的线粒体过滤器可能会无意中消除具有高代谢活性的健康细胞。此外,大多数将PCTMT与细胞质量联系起来的研究是在健康而不是患病的组织上进行的,而恶性组织通常由于线粒体DNA(MTDNA)拷贝数升高而导致的线粒体计数较高,或者表现出更高的线粒体计数[18]或MTOR途径的激活[18] [199,20]。因此,在癌症研究中,使用基于整个细胞群体的预定义阈值或中值绝对偏差来过滤较高的PCTMT细胞,可能会无意中消除恶性细胞的罕见,功能上重要的亚群。

当代小儿种群中有氧健身的参考值
摘要众多研究表明,体育活动有助于词汇整合(即“制定效应”),表明行动可以提高记忆力的性能并支持语言编码。这种现象最近被描述为“体现的学习”,或者涉及与学习内容直接相关的自我执行或自我生成的动作的学习。体现的语义认为,认知基于源自人类经验的多模式表示,并且运动过程在语言处理中起着至关重要的作用。这一证据的许多证据在于神经影像学研究表明,在发育和成人期间,无论是在词汇加工过程中招募了感觉和运动系统。对成人第二语言(L2)学习者的研究通常表明,感觉运动网络也参与L2处理,但比L1少。与L1获取相比,这可能是由于L2学习经常被脱皮的。最近,人们对促进体现学习及其对L2学习的影响的神经认知过程的兴趣越来越大。在本章中,我们回顾了由行为和神经认知研究报告的主要结果,探讨了本地语言处理和成人L2学习者中体现的语言处理和学习。1。体现的语义是找到我们如何将概念与语言标签联系起来的关键,这是我们理解我们如何获得第一语言,后来在生活中学习第二种语言的基础(另请参见Tokowicz&Tkacikova,本卷)。尽管进行了数十年的研究,但关于人脑如何将声学信号(例如[g ɪˈ tː])与特定概念(例如吉他)联系起来(Saussure,1916; Shapiro,2011年)仍然几乎没有共识。当前有两个相反的观点,这些观点与为概念信息构建的表示类型不同。根据经典的Amodal理论,认知是一个计算过程,从感知和通过操纵心理符号来创造意义(Fodor,1998; Landauer&Dumais,1997)。通过“三明治模型”隐喻描述了这一点:感觉运动系统简单地感知信息(输入),然后产生动作(输出)(Hurley,1998)。同时,将认知夹在两者之间,以便1)将感知的输入转换为Amodal符号,并将其链接到我们语义内存中的相关信息,并在我们的语义内存中链接到2)对这些符号进行操作以进行输出。本质上,知识存储在一个孤立的语义记忆系统中,独立于感觉运动过程。经典的Amodal理论并不能说明我们如何理解这些符号的现实世界含义,而这些符号本身是由其他符号定义的。挑战传统认知研究的一些基本信念,体现了理论,规定概念符号在某些时候必须与现实世界有关,并以感觉运动体验为基础(Hauk&Tschentscher,2013年)。这也可能适用于更抽象的概念,例如自由,至少在最初与个人经验相关(例如,一个孩子从请子弹中提取自己并听到“您是免费的!”)。根据体现的语义,概念表示受到感觉运动过程的影响很大,语言形式基于我们人体的感知和行动计划体系(Barsalou,1999)。其背后的关键概念之一是“相关学习原理”,根据该原则,动作感知和意义的同时存在导致神经元的共同触发,形成神经连接或分布式神经网络,以进行语义处理(Pulvermu ller,1999; 2013; 2013; 2013; 2013; 2013; 2013; 2013)。简而言之,“什么共同开火,将电线一起开火”(Hebb,1949年),例如,如果孩子经常在踢球时听到“踢”一词,那么词典语义网络,负责处理“踢”一词的词典语义网络,以及负责处理和执行运动必要的动作的人,将成为共享的网络,将会随着时间的推移而成为共享的网络。这个想法与Amodal理论形成鲜明对比,该理论声称用于概念知识和语言的表示形式独立于身体及其经验。尽管本章中描述的许多研究所描述的,但在该领域的最新研究最初是黑色和白色的,但该领域的最新研究变得更加细微,并集中在何时以及如何体现语言上。

服用抗糖尿病药物的糖尿病种群中饮酒的新数学模型:一种最佳控制方法
Romain Mathieu,Olivier Briat,Philippe Gyan,Jean-Michel Vinassa。在电荷方案和温度的几个参数下,快速充电对三个锂离子细胞周期寿命的影响的比较。应用能量,2021,283,pp.116344。10.1016/j.apenergy.2020.116344。hal- 04087500
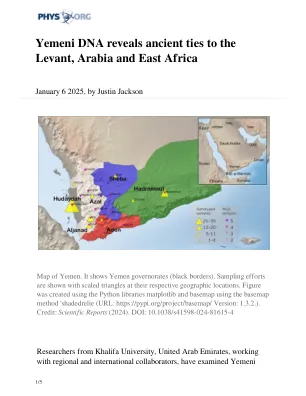
也门DNA揭示了与黎凡特,阿拉伯和东非的古老联系 AI预测药物开发受体的3D结构 合成无定形金属的新方法 欧洲人信任科学吗?新调查说“是的,但是...' “有毒男性技术”有望更快的蚊子种群生物防治 AI帮助科学家发现威胁英国湖泊生物多样性的顶级污染物 气候变化驱动蜥蜴的“生活成本”挤压 新昆虫物种突出显示科索沃生物多样性热点 AI和科学家团结起来破译由Vesuvius火山烧毁的旧卷轴 倡议要求全球合作重建过去1亿年的气氛 古代气候重建阐明了未来的海洋动力学 AI驱动的订阅模型RESHAPE零售领域,新研究找到 微藻的隐藏潜力:可持续碳捕获和行业的关键 环保墨水在3D打印中扩展了石墨烯的潜力 地球的早期周期如何塑造生命的化学 给粒子检测器一个提升:新设备的作用就像超导性开关 科学家展示了怀孕如何以无数的方式改变大脑 crispr/cas9介导的橡胶蒲公英二氨蛋白生物合成的靶向诱变 微生物会影响珊瑚漂白易感性,新研究显示 磁性纳米催化剂通过电子密度调节增强肿瘤处理 新芯片以轻速打开AI计算的门 无权手动,更安全的DNA处理的语音激活系统
沿海地区在这项研究中表现出更强的非洲混合物,而北部也门也门地区的北部地区表明与阿拉伯和黎凡特有更紧密的遗传关系。在也门漫长而持续的内战中,这项研究发现,沿海和内陆分裂的历史基因组起源不同,这与当前冲突的划分线相处。
“有毒男性技术”有望更快的蚊子种群生物防治
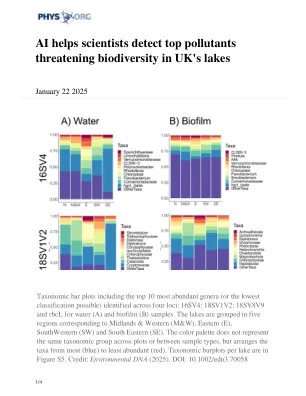
分类条形图,包括在四个基因座:16SV4上确定的前10个最丰富的属(或最低分类); 18SV1V2; 18SV8V9和RBCL用于水(A)和生物膜(B)样品。湖泊分为五个区域,与中部地区和西部(M&W),东部(E),西南(SW)和东南(SE)相对应。调色板不代表各个地块或样本类型之间的分类组,而是将大多数(蓝色)到最少(红色)的分类单元安排。每个湖泊的分类小号在图S5中。信用:环境DNA(2025)。doi:10.1002/edn3.70058

使用种群生存能力分析和化石的研究...
摘要:种群下降和灭绝通常是由多种压力源驱动的。自公元1500年以来,预测的鸟类的全球灭绝率估计比长期背景平均水平至少高出80倍。pāteke/棕色蓝绿色(Anas Chlorotis)是新西兰Aotearoa的威胁性水禽,目前的人口为c。 2500,分布在两个残余人群和少数重新引入地点。自人类到来以来,帕特克的下降是由于栖息地的丧失和破碎,捕食和其他人为相互作用而导致的。两个残余人群之一位于Aotea大屏障岛上,自1980年代以来一直在下降。我们使用了种群生存能力分析和物种分布建模的组合来更好地理解(1)AOTEA下降的驱动因素,(2)最有可能降低灭绝风险的管理干预措施以及(3)Pāteke在Aotearoa跨Aotearoa的史前分布。我们的模型通过了来自AOTEA的七年密集监控数据以及全国分布的化石记录的结合。人口生存能力分析结果表明,在接下来的100年中,AOTEA上的Pāteke人口灭绝的可能性为46%,有99%的机会降至50个人的丰度低于50个人。管理应主要关注成人的生命阶段,因为保护这个阶段导致人口增长率最大。物种分布建模结果表明,从历史上看,帕特克在沿海的大部分Aotearoa中都存在。正如人口下降通常是多种压力源的结果一样,通常需要进行多种干预措施才能停止灭绝。对于Pāteke,这将意味着控制多种哺乳动物捕食者物种,改善栖息地质量,并在其前范围内重新建立人口。对于Pāteke,这将意味着控制多种哺乳动物捕食者物种,改善栖息地质量,并在其前范围内重新建立人口。

树木种植园对西高止生物多样性热点中流行蜥蜴的种群的影响和生态学的栖息
。cc-by 4.0国际许可(未经Peer Review尚未获得认证)是作者/资助者,他已授予Biorxiv的许可证,以永久显示预印本。它是制作

成年心脏移植患者的他克莫司种群药代动力学
通讯作者:Jean-Baptiste Woillard标题:Pharmd,Phd OrcID:0000-0003-1695-0695; 0000-0001-6147-9201地址:大学。LIMOGES,INSERM U1248 P&T,2 Rue du Pr Descottes,F-87000 Limoges,法国。电话:+33 5 55 05 61 40传真:+33 5 55 05 61 62电子邮件:Jean-Baptiste.woillard@unilim.fr作者确认本文的主要研究人员是Pierre Marquet教授和

通过遗传多样性和种群结构来优化早期培养白玉米的繁殖策略
玉米的生产和撒哈拉以南非洲的生产力受到各种因素的约束。评估新开发的精英亲属线的遗传多样性可以帮助识别具有理想基因的线条并探索杂种育种的遗传相关性。这项研究的目标是评估遗传多样性和种群结构的水平,并确定适当的聚类方法,以将玉米含量分配为杂种群体。使用多样性阵列技术(DARTTAG)中密度平台对从三个来源种群中提取的三百七十六个精英杂种进行了基因分型。从1904年获得的3,305个SNP标记的结果显示,平均标记物多态性信息含量(PIC)为0.39,观察到0.02的杂合性,基因多样性为0.37,次要等位基因频率为0.29,Shannon和Simpson Intices,分别为6.86和949.09,分别为6.86和949.09,以及787.70.70.70.70.70.70.70.70.70.70.70.70。最佳亚群是由基于混合的模型和主成分分析定义的三个。平均遗传距离为0.303,从0.03(TZEI 2772×TZEI 2761)到0.372(TZEI 2273×TZEI 2832)。对于376个精英杂交的认可杂质分类,使用IBS距离矩阵和平均链接聚类方法提供了最高的辅助相关系数(0.97)。使用IBS距离鉴定了三个杂种组(HG),而Hg 1的平均连锁聚类方法具有188个近交,Hg 2具有137个,Hg 3具有59个近百列。基于血统的系统发育树与确定的异质基团表现出很大的一致性。基于潜在人口结构的F统计量显示,亚种群之间的差异为10%,遗传分化水平中等的亚群中的差异为90%(0.10)。精英杂交线表现出高度的遗传多样性,这可能有益于开发新的,早期培养的白色杂种,以减轻撒哈拉以南非洲的生产约束。

增强了全球侵入性埃及埃及种群的Zika病毒易感性
Aubry,F。等。(2020)增强了全球侵入性埃及埃及种群的寨卡病毒易感性。Science,370(6519),pp。991-996。(doi:10.1126/science.abd3663)。这是作者的最终接受版本。此版本和已发布的版本之间可能存在差异。,如果您想从中引用出版商的版本,建议您咨询出版商的版本。