XiaoMi-AI文件搜索系统
World File Search System纺锤体

PICH 通过其 DNA 转位酶和 SUMO 相互作用活动影响纺锤体组装检查点
抑制或稳定有丝分裂中的 SUMO 化都会导致染色体分离缺陷,这表明蛋白质的动态有丝分裂 SUMO 化对于维持基因组的完整性至关重要。Polo 样激酶 1 - 相互作用检查点解旋酶 (PICH) 是一种有丝分裂染色质重塑酶,它通过三个 SUMO 相互作用基序 (SIM) 与 SUMO 化的染色体蛋白相互作用,以控制它们与染色体的结合。使用条件性 PICH 耗竭/PICH 替换的细胞系,我们发现有丝分裂缺陷与 PICH 对 SUMO 化染色体蛋白的功能受损有关。PICH 的重塑活性或 SIM 缺陷会延迟有丝分裂进程,这是由纺锤体组装检查点 (SAC) 激活引起的,这由着丝粒处 Mad1 焦点的持续时间延长所表明。通过对染色体 SUMO 化蛋白(其丰度受 PICH 活性控制)进行蛋白质组学分析,确定了可解释 SAC 激活表型的候选蛋白。在已确定的候选蛋白中,PICH 缺失时 Bub1 着丝粒丰度会增加。我们的研究结果证明了 PICH 和 SAC 之间的新关系,其中 PICH 直接或间接影响着丝粒上的 Bub1 关联,并影响 SAC 活性以控制有丝分裂。

蛋白磷酸酶 1 与 Bud14 结合抑制酿酒酵母的有丝分裂退出
摘要 芽殖酵母的有丝分裂退出取决于有丝分裂纺锤体沿细胞极性轴的正确定位。当纺锤体无法准确定位时,一种名为纺锤体位置检查点 (SPOC) 的监视机制会阻止细胞退出有丝分裂。具有缺陷 SPOC 的突变体会变成多核并失去其基因组完整性。然而,对 SPOC 机制的全面了解尚不足。在本研究中,我们确定了 1 型蛋白磷酸酶 Glc7 与其调节蛋白 Bud14 相关联,这是一种新的检查点成分。我们进一步表明,Glc7-Bud14 促进了 SPOC 效应蛋白 Bfa1 的去磷酸化。我们的结果表明,两种机制并行作用以产生强大的检查点反应:首先,SPOC 激酶 Kin4 将 Bfa1 与抑制激酶 Cdc5 隔离开来,其次,Glc7-Bud14 使 Bfa1 去磷酸化以完全激活检查点效应物。

CLK 激酶抑制剂 1C8 和 GPS167 对上皮-间质转化和抗病毒免疫反应的影响揭示了它们的抗癌潜力
二杂芳酰胺基化合物 1C8 和氨基噻唑酰胺相关化合物 GPS167 可抑制 CLK 激酶,并影响多种癌细胞系的增殖。之前使用 GPS167 进行的化学基因组学筛选表明,与有丝分裂纺锤体组装相关的成分的消耗会改变对 GPS167 的敏感性。在这里,使用 1C8 进行的类似筛选也确定了参与有丝分裂纺锤体组装的成分的影响。因此,用 1C8 和 GPS167 处理的细胞的转录组分析表明,编码有丝分裂纺锤体组装成分的转录物的表达和 RNA 剪接受到影响。通过显示影响有丝分裂纺锤体组装的药物的亚毒性浓度会增加对 GPS167 的敏感性,证实了微管连接的功能相关性。1C8 和 GPS167 影响与肿瘤进展相关的通路中转录本的表达和剪接,包括 MYC 靶标和上皮间质转化 (EMT)。最后,1C8 和 GPS167 改变了参与抗病毒免疫反应的转录本的表达和可变剪接。与此观察结果一致,消耗双链 RNA 传感器 DHX33 可抑制 GPS167 介导的 HCT116 细胞细胞毒性。我们的研究揭示了 1C8 和 GPS167 影响癌细胞增殖以及转移关键过程的分子机制。

未经训练运动员肌肉酸痛发作延迟的诱发因素:一项横断面研究
DOMS是肌肉纺锤体中神经末端的急性神经元压缩轴突病[6]。它可能是从肌肉纺锤体引发的,还可能是由“封闭栅极”的催眠状态引起的,这是由微型I型I型感觉纤维增强引起的,此外,除了初始交感神经系统(SNS)抑制外,还可以在同心运动过程中保持闭合。这会导致非药理神经性疼痛[7]。doms可以作为重复偏心收缩的安全功能,因为它在肌肉纺锤体传入的感觉和运动神经元末端的微小损伤时可以解决。DOM的感觉主要由IV组传入纤维以及对各种刺激(包括化学,机械和热反应)的多模态反应。根据有效性的顺序,在IV组肌肉纤维中引起作用电位的化学物质是缓激肽,5-羟色胺,组胺和钾[8]。

使用CRISPR
染色体隔离需要在动型蛋白复合物和有丝分裂纺锤体之间进行协调,这对于两个子细胞之间的遗传分裂至关重要。动力学是一种蛋白质复合物,位于姐妹染色单体的丝粒上。在有丝分裂过程中,观察到的动物学实际上将姐妹染色质朝着用有丝分裂纺锤体的指南伸向细胞的相反两极。有人提出,stu1是一种小动物络合物中的小蛋白,有助于延迟酿酒酵母的萌芽酵母中的后期,直到每个染色体都附着在有丝分裂的纺锤体上。也有人建议Stu1与纺锤体相互作用,并在拉长时同步移动。已经提出,磷酸化可以调节Stu1的功能,并且熔体是其他动力学蛋白中已知的磷酸化位点,因此,在称为sTu1上的称为熔融基序的磷酸化位点上除去苏氨酸氨基酸在Stu1上的磷酸化位点可能会影响姐妹染色体的能力,这可能会导致姐姐的正确性,这可能会使YEAST YEAST降低。熔体是真菌中保存良好的序列,是其他动力学蛋白中的已知磷酸化位点,是STU1的同源物。利用CRISPR-CAS9酶,我们将在发芽的酵母菌Stu1基因中引入磷酸无效突变,以用熔体序列替代苏氨酸719密码子。到目前为止,我们已经成功克隆了含有引导RNA和Cas9酶基因的质粒。我们假设该突变将在Stu1中产生故障,这可能会阻碍其协调纺锤体和动孔附着的能力,并在有丝分裂过程中完全防止染色体分离。下一步将是用质粒和我们的模板DNA转化酵母,该模板DNA代码在Stu1中的719密码子上编码Valine,这种组合将完全激活酵母中的CRISPR CAS CAS 9基因组编辑系统。

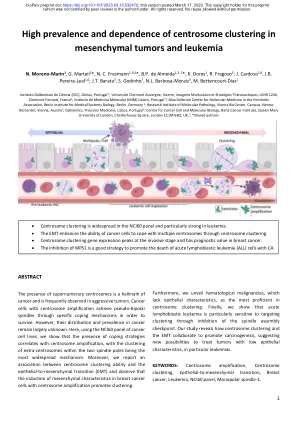
间充质肿瘤和白血病中着丝粒聚集的高患病率和依赖性
多余着丝粒的存在是癌症的一个标志,在侵袭性肿瘤中经常观察到。着丝粒扩增的癌细胞通过特定的应对机制实现伪双极纺锤体以求生存。然而,它们在癌症中的分布和流行程度仍然很大程度上未知。在这里,我们使用 NCI60 组癌细胞系,表明应对策略的存在与着丝粒扩增相关,其中两个纺锤体极内额外着丝粒的聚集是最普遍的机制。此外,我们报告了着丝粒聚集能力与上皮-间质转化 (EMT) 之间的关联,并观察到着丝粒扩增的乳腺癌细胞中间质特征的诱导促进了聚集。

生命科学
•染色质网络凝结,螺纹变短,更厚,可见为染色体。•同源染色体成对排列,以使每对的两个染色体并排躺在交叉过程中形成二价。•每个同源染色体都可以看到,因为两种染色单体由丝粒连接。单个染色体的这些相同的染色单体称为姐妹染色质被。•交叉发生在同源染色体之间。同源染色体并排躺在。他们的非姐姐染色质酸重叠并在称为chiasmata(单数:chiasma)的点上触摸。染色单体片段分解并在chiasmata上交换。以这种方式,遗传物质在同源染色体之间交换以形成重组染色质被。•跨越跨性别物质和父亲染色体之间的遗传物质通过改组或重组来引入遗传变异。•核仁和核膜消失。•在动物细胞中,中心体分裂和成对的中心元素移至细胞的相对极。•纺锤体纤维之间会在中心纤维之间形成纺锤体。

生命科学
•染色质网络凝结,螺纹变短,更厚,可见为染色体。•同源染色体成对排列,以使每对的两个染色体并排躺在交叉过程中形成二价。•每个同源染色体都可以看到,因为两种染色单体由丝粒连接。单个染色体的这些相同的染色单体称为姐妹染色质被。•交叉发生在同源染色体之间。同源染色体并排躺在。他们的非姐姐染色质酸重叠并在称为chiasmata(单数:chiasma)的点上触摸。染色单体片段分解并在chiasmata上交换。以这种方式,遗传物质在同源染色体之间交换以形成重组染色质被。•跨越跨性别物质和父亲染色体之间的遗传物质通过改组或重组来引入遗传变异。•核仁和核膜消失。•在动物细胞中,中心体分裂和成对的中心元素移至细胞的相对极。•纺锤体纤维之间会在中心纤维之间形成纺锤体。

自然界的边缘计算
像肌肉纺锤体的多个本体感受信号一样,被认为可以对身体构型进行强大的估计。然而,尚不清楚主轴信号是否足以区分肢体运动。在这里,一个模拟的4- musculotendon,2关节平面肢体模型在向前和反向方向上产生了五个终点轨迹的重复循环,从每个musculotendon产生了速度和II传入信号(分别为速度和长度)产生纺锤体和II传入信号。我们发现,原始射击率的8D时间序列的互相关(四个IA,四个II)无法区分大多数运动对(〜29%精度)。但是,将这些信号投射到其1 st和2 nd主组件上大大提高了运动对的可区分性(精度为82%)。我们得出的结论是,肌肉本体受体的高维团可以区分肢体运动,但仅在降低维度后。这可以解释到达体感皮质之前的某些传入信号的预处理,例如在猫的cuneate核上处理皮肤信号。