XiaoMi-AI文件搜索系统
World File Search System组可

Gal-1 和Gal-9 在子痫前期患者血清和胎盘表达的临床研究
0.05), 且早发型 PE 组 Gal-1 水平和 Gal-9 水平亦显着高于晚发型 PE 组 ( P <0.05)。 早发型 PE 组和晚发型 PE

2025年生物信息学与计算生物学国际学术会议
基因组学和疾病研究、高通量数据分析、网络生物学、计算遗传学、模型解释和可视 化、生物数据挖掘、比较基因组学、机器学习和医学影像分析、蛋白质结构与功能预测、 宏基因组学与微生物组、知识图谱构建、生物信息学工具开发、转录组学和表达谱的分析、 药物发现与设计、遗传流行病学、蛋白质组学、个性化医疗与精准医学、生物医学工程、 结构生物信息学、计算工具和软件开发、进化生物信息学、系统生物学、环境与生态计算 生物学和流行病学、计算生态学、序列分析、模式识别与生物信号处理、生物信息学与统 计分析、下一代测序技术、计算生物学与人工智能的融合、生物数据挖掘、处理与分析、 计算医学与临床应用、代谢组学、生物信息学工具与网络科学。

无人机系统 (UAS) 第 1 组和第 2 组物理...
无人机系统 (UAS) 第 1 组和第 2 组体检工作表由合格的医疗服务提供者检查。任何不合格情况或“不合格”部分都需要在第 8 块中注明并由医疗官审查。向成员的指挥官 (CO) 提交豁免请求。参见 MANMED CH-15 第 IV 节
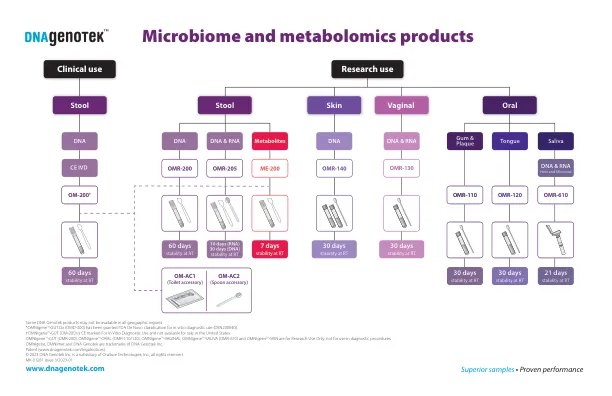
微生物组和代谢组学产品
在所有地理区域中,某些DNA Genotek产品可能不可用。†Omnigene TM•肠道(OM-200)标记为体外诊断使用,在美国不可出售。Omnigene TM•肠道(OMR-200),Omnigene TM•口服(OMR-110/120),Omnigene TM•阴道,综合TM•Saliva(OMR-610)和Omnigene TM•皮肤仅用于研究,不适用于诊断过程。Omnigene,Omnimet和DNA Genotek是DNA Genotek Inc.专利的商标(www.dnagenotek.com/legalnotices)©2023 DNA Genotek Inc.是Orasure Technologies,Inc.,Inc.,Inc。,保留所有权利。MK-01281第3/2023-01号

集成的转录组和代谢组分析...
©作者2025。Open Access本文在创意共享属性下获得许可 - 非商业 - 非洲毒素4.0国际许可证,该许可允许以任何中等或格式的任何非商业用途,共享,分发和复制,只要您与原始作者提供适当的信誉,并为您提供了符合创造性共识许可的链接,并提供了持有货物的启动材料。您没有根据本许可证的许可来共享本文或部分内容的适用材料。本文中的图像或其他第三方材料包含在文章的创意共享许可中,除非在信用额度中另有说明。如果本文的创意共享许可中未包含材料,并且您的预期用途不受法定法规的允许或超过允许的用途,则您需要直接从版权所有者那里获得许可。要查看此许可证的副本,请访问h t p://c r e a t i v e c o m m o ns。or g/l i c e n s e s/b y-n c-n c-n c-n d/4。0/。
肠道微生物组和代谢组签名...
研究亮点•肠道微生物营养不良与肝硬化进展显着加剧,这是特定微生物和代谢产物的同时发生的变化。有趣的是,肝硬化诱导的肠道和代谢产物的转移增加与与肝硬化相关的临床标记密切相关。这项研究扩大了针对特定肝硬化相关并发症的肝硬化生物标志物的范围。此外,这项研究高点肝硬化患者的粪便微生物和代谢标志物的相关性与临床标志物紧密相关,例如MELD和CTP评分,以及AST,ALT,ALT,ALT,胆红素和γ-GT水平。这些发现增强了我们对肝硬化中肠道微生物组的理解及其与相关的微生物物种和代谢产物的联系。

枳果梗多糖调节谷氨酸代谢和紧密连接蛋白表达改善酒精 ...
摘 要 : 目的:本研究旨在明确枳椇果梗多糖( HDPs )对酒精暴露所致的小鼠神经行为异常的改善效果,并探究谷 氨酸代谢和紧密连接蛋白表达在其中的作用。方法:雄性 C57BL/6 小鼠按 114 μL/20 g 剂量连续酒精灌胃 14 d ,建 立酒精暴露模型,同时设置干预组进行 HDPs 干预( 114 μL/20 g 酒精 +100 mg/kg HDPs )。应用行为学实验(旷场 实验、高架十字迷宫实验)评估神经行为学变化,采用气相色谱法测定小鼠血液中乙醇浓度, γ -H2AX 荧光检测小 鼠脑海马组织 DNA 损伤,免疫组化分析检测小鼠脑组织中紧密连接蛋白 Claudin-1 和 ZO-1 的表达,并通过超高 效液相色谱 - 四级杆飞行时间质谱法( UPLC-Q-TOF-MS )代谢组学技术对小鼠脑组织代谢物进行分析。结果: HDPs 可有效降低酒精暴露小鼠血液乙醇浓度,由 4.69±0.29 g/L 降至 1.64±0.104 g/L ;改善酒精暴露所致的小鼠神 经行为异常,旷场实验中,与酒精组相比, HDPs 干预组总路程显着提升至 27340±3304 cm ( P <0.05 ),平均速度 显着提升至 67.4±13.4 cm/s ( P <0.05 ),不动时间缩短 29% ( P <0.05 );高架十字迷宫实验中,与酒精组相比, HDPs 干预组闭臂停留时间显着减少至 195.6±10.3 s ( P <0.05 ),开放臂进入次数显着增加 26% ( P <0.05 ));还 可降低酒精诱导的脑组织氧化应激与 DNA 损伤水平, ROS 、 MDA 分别降低 5.4% 、 29.5% ( P <0.05 ), T-AOC 提 高 10.9% ,上调脑海马组织中 Claudin-1 ( 2.2 倍)和 ZO-1 ( 0.1 倍)蛋白的表达;并调节脑组织谷氨酸代谢通路, 提高甘氨酸( 19.7% )、谷光甘肽( 25% )、琥珀酸( 22.6% )等代谢物水平。结论: HDPs 可有效改善酒精对小鼠 神经行为的影响,其机制或可能通过抗氧化、保护紧密连接蛋白和调节谷氨酸代谢通路发挥作用,研究结果可为 扩展枳椇资源在食品领域中的应用提供理论依据。

组异质性
尽管有几种备受瞩目的最先进的方法可用,但分析批量RNA-Seq数据仍在面临重大挑战。最近的研究的证据表明,流行的差异表达(DE)工具(例如EDGER和DESEQ2)容易受到惊人的错误发现率(FDR)的影响。这些研究表明,在这些模型中观察到的FDR通货膨胀可能归因于诸如违反参数假设的问题或无法有效处理数据中的异常值。在这里,我们认为群体异质性也可以促进这一提升的FDR,这一现象在很大程度上被研究界忽略了。我们介绍了一种新型的统计模型Robseq,该模型旨在在差异分析中有效的每种功能建模,当时是当群均均一的假设未得到满足时。Robseq利用了稳健的统计文献中建立的统计机制,包括M估计量来稳健地估计基因表达水平变化和Huber-Cameron方差估计器来计算异性设置中的鲁棒标准误差。此外,出于推理目的,它还结合了Welch T统计量的自由度调整,有效地解决了RNA-Seq差异表达中FDR通胀的问题。通过详细的模拟和全面的基准测试,我们表明Robseq成功地将错误的发现和I型错误率保持在名义级别,同时与众所周知的DE方法相比保留了高统计能力。对种群级RNA-seq数据的分析进一步表明,Robseq能够鉴定出与复杂人类疾病有关的具有生物学上重要的信号和途径,这些信号和途径涉及复杂的人类疾病,否则这些信号和途径否则无法通过已发表的方法揭示。Robseq的实现可在https://github.com/schatterjee30/robseq上公开提供。

