XiaoMi-AI文件搜索系统
World File Search System细胞核
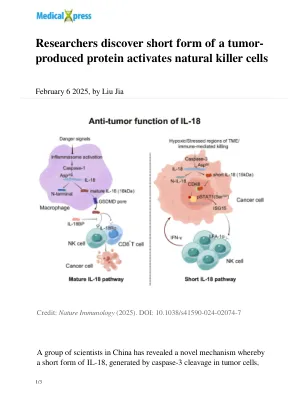
产生的蛋白质激活天然杀伤细胞
在这项研究中,研究人员专注于IL-18。他们发现肿瘤细胞可以通过caspase-3裂解产生新型的IL-18形式,该裂解与传统的成熟IL-18途径无关。与成熟的IL-18不同,这种短形式不会退出细胞,而是进入细胞核,在该细胞核中促进了STAT1和ISG15分泌的磷酸化,从而增强了NK细胞抗肿瘤功能。

细胞骨架成分的先天突变引起DNA dna甲基化...
uhrf1在受精后主要迁移到卵和胚胎中的细胞质,其中少量的UHRF1在某些区域(例如ICR)中维持甲基化修饰的细胞核中剩余少量。另一方面,除了受精后立即卵和胚胎外,所有UHRF1均易位到细胞核中,并在与细胞分裂相关的DNA复制过程中复制甲基化修饰。由于使用卵的实验受到局限性,因此研究小组使用人类培养的细胞发现NLRP5和OOEEP与构成SCMC的核心蛋白之间的结合。研究小组还产生了一条细胞系,可以通过药物诱导的诱导UHRF1(称为Cuhrf1:图1),该细胞系已被修饰以将其定位为细胞质,就像卵子一样,并检查了Cuhrf1在NLRP5和OOEP存在下CuHRF1变化的蛋白质稳定性。我们发现,在OOEEP存在下,CuHRF1的稳定性不会改变,但是在NLRP5存在下,Cuhrf1的稳定性增加了两倍以上(图2)。我们还发现,NLRP5缺陷小鼠的卵中的细胞质和细胞核中UHRF1蛋白的量均降低。该结果表明,在易位进入细胞核后,稳定的UHRF1的一部分可能稳定存在。

胰岛素调节有丝分裂原激活的蛋白激酶激酶(MEK),有丝分裂原激活的蛋白激酶和酪蛋白激酶在细胞核中:Possibl
在大鼠大脑皮层中研究了腺苷酸环化酶和鸟嘌呤核苷酸结合蛋白(G蛋白)在锂对脑功能的慢性作用中的可能作用。发现,用锂(具有治疗相关的血清水平为1 mm)对大鼠的慢性治疗增加了mRNA和蛋白质的水平,用于钙调蛋白敏感(1型)和钙调蛋白敏感(2型)形式的腺苷酸环化酶和抑制蛋白质的mRNA和蛋白质水平降低,用于抑制性gja2 gja2 gja2 gja2 gja2 gja2。慢性锂不会改变其他G-蛋白亚基的水平,包括GA,GSA和GJF。在短期锂治疗(最终血清水平为-1 mM)或以较低剂量的锂(血清水平为-0.5 mm)下,h含腺苷酸环化酶和GIA的锂调节均未观察到短期锂治疗(最终血清水平为-1 mm)。结果表明,腺苷酸环化酶的上调和GJA的下调可能代表了分子机制的一部分,锂可以改变脑功能并在治疗情感障碍的治疗中发挥其临床作用。

囊性纤维化:DNA 案例研究
遗传结构对话快照 生物课后吃午饭时,四个朋友讨论 DNA、基因和染色体。这是对话快照。 Billy 说“我对 Henderson 女士今天在课堂上告诉我们的内容有点困惑。DNA 存在于细胞核和基因中。基因组成染色体,对吗?” “我听到她说的不是这个。我认为染色体组成 DNA,而 DNA 存在于基因上。我同意你关于整个细胞核的事情,”Kyle 说。 “得了吧伙计们,”Kelly 说“她根本不是这么说的。她告诉我们基因是 DNA 的一部分,而 DNA 组成染色体。” Janet 说,“Kelly,你搞反了。染色体存在于基因的 DNA 上。我不确定整个细胞核的事情,但我确信所有这些都存在于细胞中。” 使用右侧的空白处回答以下问题。你可以画图来帮助说明你的回答。您认为谁对 DNA、染色体和基因的理解最透彻?为什么?

通过核肌动蛋白
摘要越来越多地赞赏,核的结构成分通过改变染色质组织来调节基因可及性。虽然核膜连接器蛋白将机械敏感性肌动蛋白细胞骨架与核骨架联系起来,但肌动蛋白对核内部结构的贡献仍然神秘。控制肌动蛋白转运到细胞核中,加上控制肌动蛋白结构(肌动蛋白工具盒)的蛋白质的存在,这表明核肌动蛋白可以支持基因表达的生物力学调节。细胞肌动蛋白结构是机械响应性的:通过在质膜传播力在细胞核中传播的力产生的肌动蛋白电缆。我们认为,对这种生物力学提示的响应动态肌动蛋白重塑为表观遗传景观提供了新的结构控制水平。我们在这里提出要对机械力可以促进肌动蛋白转移到细胞核和控制结构排列的事实中,如间充质干细胞中所示,从而调节谱系承诺。



误解:COVID-19 疫苗不安全,因为它开发得太快了。事实:疫苗已被证明是安全有效的。尽管它们
谣言:mRNA COVID-19 疫苗会改变我的 DNA。事实:没有疫苗会改变你的 DNA。这些类型的疫苗含有信使 RNA (mRNA),它指示细胞制造新冠病毒上的“刺突蛋白”。当免疫系统识别出这种蛋白质时,它会通过产生抗体来建立免疫反应,从而教会身体如何预防未来的感染。mRNA 永远不会进入细胞核,而细胞核正是我们 DNA 的保存地。身体在完成指令后会清除 mRNA。

mRNA 技术 - 洞察报告
mRNA 在转录过程中在细胞核中合成,产生前 mRNA,然后加工成 mRNA。在转录过程中,遗传信息由 RNA 聚合酶从 DNA 中复制,形成所谓的前 mRNA。然后,该分子通过添加 5' 帽和 3' poly(A) 尾巴进行加工,并通过在细胞核中剪接内含子序列形成五组分成熟 mRNA 结构。mRNA 结构中的每个组分在细胞质中核糖体的运输、翻译和有效生产蛋白质方面都有特定的作用(见图 3)。

弓形虫线粒体转运蛋白 ABCB7L 对细胞浆和细胞核铁硫簇蛋白的生物合成以及细胞浆翻译至关重要
摘要 铁硫 (Fe-S) 簇是普遍存在的无机辅因子,是许多细胞必需途径所必需的。由于它们不能从环境中清除,因此 Fe-S 簇在细胞区室(如顶质体、线粒体和细胞质)中从头合成。细胞质 Fe-S 簇生物合成途径依赖于线粒体途径中间体的运输。一种称为 ABCB7 的 ATP 结合盒 (ABC) 转运蛋白在许多常见研究的生物体中负责这一作用,但它在医学上重要的顶复门寄生虫中的作用尚未被研究。在这里,我们识别并描述了一种弓形虫 ABCB7 同源物,我们将其命名为 ABCB7-like (ABCB7L)。基因耗竭表明它对寄生虫的生长至关重要,并且它的破坏会触发部分阶段转换。敲除系的表征突出了细胞质和细胞核 Fe-S 蛋白的生物合成缺陷,导致蛋白质翻译和其他途径(包括 DNA 和 RNA 复制和代谢)出现缺陷。我们的工作为广泛保留 Fe-S 簇生物合成中线粒体和细胞质途径之间的联系提供了支持,并揭示了其对寄生虫生存的重要性。