XiaoMi-AI文件搜索系统
World File Search System编码
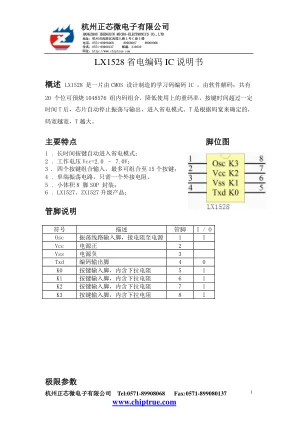
LX1528 省电编码IC 说明书
HANGZHOU ZHENGXIN MICRO-ELECTRONICS CO.,LTD 地址:杭州市西湖区西园八路 2 号 C 座 3 楼 电话: 0571-89908068 89908067 89908066 传真: 0571-89908137 邮编: 310030 网址: http://www.chiptrue.com 符号 参数 条件 范围 单位 Vcc 电源电压 -0.3 ~ 7 V VI 输入电压 -0.3 ~ Vcc+ 0.3 V VO 输出电压 -0.3 ~ Vcc+ 0.3 V Tst 储存温度 -40 ~ 125 ℃ Top 工作温度 - 20 ~ 70 ℃ Pdis 最大功耗 Vcc = 6V (空载) 10 mW

长链非编码RNA 编码的微肽研究进展
[摘要]长的非编码RNA(LNCRNA)是由200多个核苷酸构成的RNA分子,表现出相对较低的序列保护。很长一段时间以来,它们被视为“转录噪声”,即在生物领域中的非功能性RNA分子。近年来,随着研究的进步,科学家们在lncrnas中揭示了许多小型开放式阅读框(SORF),其中一些可以编码微肽。这些微肽已被证实参与了各种细胞过程和基因表达调节网络,扮演着至关重要的作用。这一发现为进一步探索生活活动以及临床诊断和疾病治疗的新研究方向开辟了新的研究方向。本综述总结了LNCRNA编码的菌根在病理和生理过程中的作用,微肽的亚细胞定位和功能机制以及微肽研究方法的进展,旨在为新型积分基于磨性的诊断诊断和治疗方法提供洞察力和参考。[关键词]长的非编码RNA;小开放阅读框;微肽;肿瘤

10 编码
方法:本研究分析了马来西亚卫生信息中心 2017 年 1 月至 2022 年 6 月的死亡记录,这些记录已编入 ICD-10。数据匿名化符合道德标准,经过质量检查后,共纳入 387,650 份死亡登记。数据集仅限于三位数的 ICD-10 代码,经过清理和 80:20 的训练测试分割。预处理涉及 HTML 标签删除和标记化。ML 方法,包括 BERT(来自 Transformer 的双向编码器表示)、Gzip+KNN(K 最近邻)、XGBoost(极端梯度提升)、TensorFlow、SVM(支持向量机)和朴素贝叶斯,都已针对自动 ICD-10 编码进行了评估。使用 Amazon SageMaker(亚马逊网络服务,华盛顿州西雅图)对模型的准确度、F1 分数、精确度、召回率、特异性和精确度-召回率曲线进行了微调和评估。敏感性分析解决了不平衡数据场景,增强了模型稳健性。

rachel-claire-farnsworth-relese-new-new-book编码 -
多国赢得健康治疗师和创伤发行专家Rachel Claire Farnsworth资料来源:Rachel Farnsworth



什么是编码失败
encoding failure occurs when receiver unable interpret data due incompatible encoding schemes this lead corruption or unreadable data cause usually different computing systems use different encoding methods encoding used represent store communicate digital information example some systems use ascii american standard code for information interchange while others use utf-8 unicode transformation format 8-bit if system attempt send information encoded one method but receiver uses different method then encoding failure occur in addition incompatible coding standards encoding failures can also caused by incorrect character sets or technical errors transmission minor discrepancies sender's receiver's coding standards can cause error fortunately several ways prevent encoding failures most effective ensure both parties use same coding standard before sending data verify all characters message correctly encoded before transmitting default coding standard use unicode accommodate almost all languages character sets Failure in Memory Retention: Causes and Consequences The failure to retain information in长期记忆可能由于各种因素而发生,包括缺乏积极参与,助记符设备的使用不佳,实践检索不足以及对其他记忆的干扰。####编码故障类型的类型有三种主要类型的编码失败:1。**编码失败**:当信息未编码为长期内存时,就会发生这种情况,从而无法进行检索。2。**存储衰减**:当信息被编码时,这会发生,但是由于神经元或它们之间的路径损坏而随着时间的流逝而衰减。3。但是,如果此过程被中断怎么办?**检索失败**:这种故障会发生,尽管编码和存储正确并存储了长期记忆,但会发生这种故障。####对编码几个因素的干扰因素可能会妨碍编码,包括:**其他记忆中的干扰**:当项目与其他记忆具有相似之处时,正确编码可能是具有挑战性的。***彩排干扰**:重复自己的头部,而不是试图记住它会使它难以保留。***认知负载**:由于多个任务或分心而导致的过多认知负荷可能会阻碍编码。#### Examples of Retrieval Failure Retrieval failure can manifest in various ways, such as: * Forgetting recent activities * Struggling to recall names or phone numbers * Difficulty accessing information from long-term memory #### Strategies for Overcoming Encoding Failures To improve encoding and retention, consider the following strategies: * Practice active engagement with the material * Utilize mnemonic devices to aid in organization and recall * Engage in regular practice retrieval to加强学习信息基于各种方法(例如它的所见,听到或含义)存储在内存中。编码和解码是将书面符号变成可理解的形式的过程。在编码中,我们使用单个声音来构建单词,而在解码时,我们大声朗读或将书面单词转换为可理解的形式。要阅读,我们将字母解码为它们相应的声音,然后在我们的脑海中构建单词,这对我们大多数人都会自动发生。自我参考效应还通过将信息与自己联系起来有助于记忆。编码有些不同,需要了解单个声音并以正确的顺序将它们放在一起。语义编码涉及将含义附加到信息上并将其连接到相关信息,从而更有效。健忘可能是由压力,抑郁,缺乏睡眠,甲状腺问题或某些药物副作用引起的。如果编码数据不正确,则可能会导致数据的显示或解释方式。可以通过将实际记忆与通过催眠收到的他人收到的建议相结合,或使用照片或其他图像来植入虚假记忆来创建错误的记忆。编码失败,一种心理现象,可能会对我们的日常生活及其他地区产生深远的影响。在当今快节奏的世界中,记忆形成在塑造我们的身份方面起着巨大的作用。这样想:当您学习新知识时,您的大脑会进行精神舞蹈来处理该信息。那是编码故障的地方 - 系统中的一个故障,使我们争先恐后地记住我们从未真正学到的东西。不喜欢忘记您已经知道的东西 - 这更像是从来没有一开始就写下来。想象一下在聚会上遇到一个新人,但他们的名字像手指之间的沙子一样从您身上滑落。那是在您眼前发生的编码失败。我们的大脑不断受到信息的轰炸,这取决于我们专注于真正重要的事情。持续编码失败可能会随着时间的流逝而导致认知能力下降。但是,当我们过于陷入多任务处理或被太多数据所淹没时,我们的大脑可能难以跟上 - 导致那些令人沮丧的遗忘时刻。即使是压力和情感上的东西也可能会阻碍 - 就像当您如此担心某些东西时甚至无法记住放置钥匙的地方。并且不要忘记身体上的因素 - 如果我们在听力或看见(例如听力或看见)中苦苦挣扎,它可能会影响我们学习新信息的程度。编码困难可能源于初始感知,神经系统条件以及影响大脑有效编码新记忆能力的各种其他因素。这可能会带来巨大的后果,影响学术环境中的学习和表现,个人生活中的关系甚至法律程序。诊断编码问题由于其微妙的性质可能是具有挑战性的,但是心理学家和神经科医生使用各种工具和技术,包括认知评估,记忆测试和神经影像学方法,例如功能磁共振成像(fMRI)。自我报告的症状和行为观察在诊断中也起着至关重要的作用。必须将编码失败与其他记忆障碍(例如存储或检索故障)区分开。幸运的是,个人可以采用一些策略来提高其编码能力,包括通过冥想或集中呼吸练习等正念技巧提高注意力和专注。编码故障可能是一个重大问题,但是采用助记符设备和记忆辅助工具(例如基因座方法)可以帮助改善心理联系和保留。生活方式的变化,例如定期运动,均衡饮食和足够的睡眠也会有助于最佳的大脑健康。努力的编码技术,例如总结信息或创建视觉表示形式可以显着改善记忆力保留。在某些情况下,可能需要采取医疗干预措施来解决严重或持续的编码问题。研究人员正在探索新的途径,包括针对记忆形成的脑部计算机界面和基因疗法。对编号和语义编码的研究也是一个激烈研究的领域,旨在开发针对编码故障的针对性干预措施。认识到编码在日常生活中的作用,了解其原因和后果,并采取主动步骤可以改善认知功能和更加联系的生活。编码需要积极的参与和努力;采用诸如详细编码之类的技术可以改善内存形成。在编码失败时对自己友善至关重要,将它们视为学习和成长的机会,而不是使自己殴打。您可以采用根据您的需求量身定制的个性化策略来增强您的编码能力。编码和记忆形成之间的复杂关系揭示了人类认知的复杂性。编码失败是一种普遍现象,但它是增长的机会。通过确认其意义,您可以采取积极的步骤来增强记忆创造的关键方面。您可以利用各种技术,例如正念实践,助记符设备或生活方式修改,以提高编码效率。研究继续提高我们对编码过程的理解,新发现使我们更加接近释放人类记忆的全部潜力。通过好奇和同情心的挑战,您可以将看起来像是一定的机会转变为与人类心理学复杂性更深入地互动的机会。

编码项目
内容本课程介绍了算法解决问题。其主要目标是学习如何通过使用最合适,最有效的数据结构来建模由管理工程引起的实际问题,以及如何通过使用经典算法和图形理论来实现最有效的解决方案方法。该课程强调了数字化以及算法与编程之间的关系的重要性,以及通过开发旨在解决每年分配的特定问题的最终编码项目,与项目管理和解决问题的技能相关的方面。该问题可能由管理工程或计算机科学的任何领域引起;它可能享受任何路由,分区,着色,位置,电信,可持续物流和供应链管理,投资组合,调度,数据挖掘或业务分析功能,并且可能具有任何一般结构。学生将必须小组工作以最有效的方式解决和解决问题,并准备在考试期间捍卫自己的工作。课程特别包括以下主题:

战术显控系统人机交互中的特征整合设计研究 - 包装工程
编码特征作为预测结果,邀请用户进行认知情况调 研。从用户调研数据的计算结果可知,用户对不同特 征编码的认知存在一定的共性,有共同的认知习惯。 1 )就属性语义来看,认知效率主要受色相、明 度、饱和度、尺寸、位置、形状的影响。色相:国军 标对色彩的应用有明确的规范,在进行色相编码时, 应考虑用户对专用色彩属性的认知习惯,严格遵守色 彩使用规范。对于没有硬性规定的色彩,也应以用户 过往的知识、经验为基础进行编码设计。如,在界面 设计中,一般认为红色表示危险,黄色表示警告,绿 色表示安全。明度:实验表明,在深色背景下,明度 越高信息等级越高。战术显控系统复杂性较高,合适 的明度编码设计适合应用于信息层级设计,能够有效 降低用户的学习成本。饱和度:饱和度取决于该色中 含色成分和消色成分(灰色)的比例。含色成分越大, 饱和度越大;消色成分越大,饱和度越小 [14] 。高饱和 度的色彩编码方式更能引起视觉关注,帮助用户集中 注意力。形状:在战术显控系统中,涉及形状属性的 元素主要为图形和符号,包括通用类和特殊类。在进 行形状编码时,现有图符应遵循沿用的原则,新的图 符应结合现实形态、行业背景进行设计,以符合用户 认知习惯、缩短学习过程,提高交互效率。尺寸:根 据实验结果显示,信息尺寸的大小与信息的重要等级 成正比,信息越重要,尺寸越大。位置:用户对显示 屏上的信息关注度依次为中间、左上方、右上方、左 下方、右下方 [15] 。在进行界面布局时,应注意信息等 级与其在界面中位置的一致性,同时要保证同类信息 的位置编码统一。 2 )就情感语义来看,战时用户的生理和心理负 荷较高,任务情景的不确定性易增加用户的操作压 力 [5] 。在进行交互界面设计时应考虑信息编码元素的 情感性。从实验结果来看,影响情感语义的特征主要 为形状和色彩。尖锐的形态容易让用户产生较大的心 理压力,而圆润浑厚的形状更容易使用户平静。在进 行形状编码时,可采用倒角的设计手法。根据蒙赛尔 色彩体系对色彩要素的划分及实验结果,战术显控系 统的主色可以选用冷色调,明度、饱和度不宜过高, 以避免色彩刺激增加用户的焦虑感。而对于重点信息 和即时变化类信息,可采用高明度或高饱和度的色 彩,以提高用户的警觉性。

计费和编码指南
参考文献 1. 美国疾病控制与预防中心。JYNNEOS 疫苗。访问日期:2024 年 1 月 10 日。https://www.cdc.gov/poxvirus/mpox/interim-considerations/jynneos-vaccine.html 2. 美国疾病控制与预防中心。ACIP 建议。访问日期:2024 年 1 月 10 日。https://www.cdc.gov/vaccines/acip/recommendations.html 3. JYNNEOS 处方信息。巴伐利亚北欧;2023 年 4. 美国疾病控制与预防中心。免疫信息系统 (IIS):HL7 标准代码集 CVX – 已接种疫苗。访问日期:2024 年 2 月 13 日。https://www2a.cdc.gov/vaccines/iis/iisstandards/vaccines.asp?rpt=cvx 5. 美国疾病控制与预防中心。免疫信息系统 (IIS):HL7 标准代码集 MVX – 疫苗制造商。访问日期:2024 年 2 月 13 日。https://www2a.cdc.gov/vaccines/iis/iisstandards/vaccines.asp?rpt=mvx 6. 美国医学会。2024 CPT 专业版。现行程序术语 (CPT ® ) 版权所有 2024 美国医学会。保留所有权利。CPT ® 是美国医学会的注册商标。伊利诺伊州芝加哥:AMA;2024。7. 北卡罗来纳州医疗补助健康福利司。猴痘疫苗 (Jynneos ™ ) HCPCS 代码 90611:计费指南。访问日期:2024 年 2 月 14 日。https://medicaid.ncdhhs.gov/blog/2022/10/07/monkeypox-vaccine-jynneostm-hcpcs-code-90611-billing-guidelines 8. ICD10Data.com。2024 年 ICD-10-CM 诊断代码 Z23。访问日期:2024 年 2 月 13 日。https://www.icd10data.com/ICD10CM/Codes/Z00-Z99/Z20-Z29/Z23-/Z23。