XiaoMi-AI文件搜索系统
World File Search System肥大细胞

开发一种评估组织活检中肥大细胞和嗜酸性粒细胞的标准化方法:AAAAI过敏性皮肤疾病委员会的AWORK组报告
来自病理学和实验室医学和C儿科的部门,P,辛辛那提大学医学院内科学系,内科学系,内科学系,B胃和免疫学和G胃肠病学,HEPTOTOLOGY和HEPTOTOLOGY和HAPTOTOLOGY and NATICTION,CINCINNATI CHIL-CHIL-DREN'S CENTER; d过敏和免疫学部,科罗拉多大学科罗拉多大学内科部门; e密歇根大学安阿伯分校内科学系过敏和免疫学系; g巴尔的摩约翰·霍普金斯医学院内科学系过敏和免疫学系; H医学院,里约热内卢医学院; l Excelencia en asma y Alergia中心,医院M Edica Sur,Ciudad de M exico; j哮喘和过敏中心,Lew-Isville; K哮喘和过敏中心,花丘; l德克萨斯大学西南医学中心的过敏和免疫学系; m盐湖城犹他大学健康科学中心医学系胃肠病学系; n胃肠病学,肝病学和营养,过敏和哮喘中心,麦克莱恩;和0芝加哥的Ann和Robert H. Lurie儿童医院,儿科学系过敏和免疫学的司。披露潜在的利益冲突:PA已从美国国立卫生研究院(NIH)/美国国家过敏和传染病研究所获得赠款(U54 AI117804和R01 AI124355-01),患者居中的现象研究所(SC14-1403-1403-11593)和SHIRE和SHIRE和SHIRE和SHIRE,ca已获得蓝图药物的研究赠款支持,是SCD已获得NIH和Genentech,Inc的赠款支持;是医学专家小组,卫生和公共服务部以及疫苗伤害补偿部的成员;并在顾问委员会和/或是Allakos,CSL Behring,Biocryst,Grifols和UKKO的顾问。

注射:药物 T 政策
气道炎症是哮喘发病机制中的一个重要组成部分。多种细胞类型(例如肥大细胞、嗜酸性粒细胞、中性粒细胞、巨噬细胞、淋巴细胞、ILC2 细胞)和介质(例如组胺、二十烷酸、白三烯、细胞因子)参与气道炎症。使用 tezepelumab-ekko 阻断 TSLP 可减少与炎症相关的生物标志物和细胞因子,包括血液嗜酸性粒细胞、气道黏膜下嗜酸性粒细胞、IgE、FeNO、IL-5 和 IL-13;然而,tezepelumab-ekko 在哮喘中的作用机制尚未明确。

波兰寄生学会第70卷
摘要生物学中的许多数据是正态分布的,t检验,差异,回归和相关性的分析以分析这些类型的数据。但是,许多寄生虫数据并未遵循正态分布,包括卵子计数,蠕虫负担,抗体反应以及嗜酸性粒细胞和肥大细胞反应的组织学计数。因此,传统的实验设计可能不足,而传统的分析方法可以给出误导性的结果。合适的分析方法包括通用线性模型,广义线性模型,混合模型和蒙特卡洛·马尔可夫链(MCMC)Pro cedures。幸运的是,可以进行这些分析的程序可以广泛且自由使用。将使用自然和故意感染的数据来说明这些程序的使用。

工程干细胞抗体配对逃避1(...
我们的主要抗 CD117 mAb mAb-7 与未编辑的 HSPC 具有高亲和力,但与 CD117 编辑的细胞不结合(图 4)。mAb-7 与 CD117 结合可阻断 SCF-CD117 相互作用(图 4),从而选择性抑制未编辑 HSPC 的存活(图 5、6)。体外用 mAb-7 处理 HSPC 导致未编辑 HSPC 活力降低 85% 以上,而 CD117 编辑的细胞不受影响(图 5、6)。由于 CD117 信号传导与肥大细胞脱颗粒有关,我们评估了天然和 Fc 工程化版本的 mAb-7 对体外培养分化的肥大细胞的影响。Fc 工程化版本的 mAb-7 在体外不会导致任何程度的肥大细胞脱颗粒(图 6)。我们在体外比较了我们修饰的 CD117 与野生型 CD117 的磷酸化(图 7)。我们的 CD117 变体蛋白正常与 SCF 结合,在 SCF 结合后经历了相似水平的磷酸化(与 WT 蛋白相比)。我们在 HSPC 中实现了约 80% 的双等位基因 CD117 编辑和近乎完全的 HBG1/2 基因座编辑(图 8)。在异种移植研究中,我们观察到 CD117 编辑的 HSPC 能够在免疫功能低下的小鼠中进行长期多谱系造血植入(图 9)。重要的是,mAb-7 治疗导致仅使用未编辑的人类 CD34+ 细胞人源化的小鼠的人类嵌合体以及骨髓 CD34+ 细胞频率显着降低。有趣的是,在接受编辑:未编辑混合物的小鼠中,mAb-7 导致 CD117 编辑细胞在整个骨髓和 CD34+ 细胞区室中富集,这由这些区室中的高编辑水平表明(图 10)。

COVID-19 阿斯利康疫苗分析 打印
病例系列药物分析打印名称:COVID-19 阿斯利康疫苗分析打印报告运行日期:2021 年 11 月 25 日数据锁定日期:2021 年 11 月 24 日 18:30:04 MedDRA 版本:MedDRA 24.1 反应名称总计致死血液疾病贫血缺乏症贫血叶酸缺乏症 2 0 贫血维生素 B12 缺乏症 15 0 缺乏性贫血 1 0 缺铁性贫血 17 0 恶性贫血 6 0 贫血 NEC 贫血 174 0 自身免疫性贫血 1 0 失血性贫血 4 0 低色素性贫血 3 0 小细胞性贫血 1 0 正细胞性贫血 4 0 贫血溶血性 NEC 溶血性贫血 10 0 贫血溶血性免疫性自身免疫性溶血性贫血 8 0 感冒型溶血性贫血 1 0 埃文斯综合征 1 0 贫血 溶血性机械因素 非典型溶血性尿毒症综合征 1 0 红细胞碎裂综合征 1 0 出血倾向 瘀伤倾向增加 137 0 自发性血肿 8 0 自发性出血 2 0 凝血因子缺乏 获得性血友病 1 0 低纤维蛋白原血症 4 0 凝血病 凝血因子异常 11 0 抗磷脂综合征 29 0 凝血病 78 0 弥漫性血管内凝血 19 2 高凝状态 24 1 高纤维蛋白原血症 1 0 低凝状态 1 0 嗜酸性粒细胞疾病 嗜酸性粒细胞增多症 22 0 嗜酸性粒细胞增多综合征 1 0 血液囊肿和息肉 脾囊肿 1 0血液系统疾病 血液疾病 14 1 骨髓疾病 2 0 骨髓水肿 1 0 肥大细胞活化综合征 10 0 肥大细胞增多症 3 0 高铁血红蛋白血症 1 0 溶血 NEC 溶血 11 0 白细胞增多症 NEC 白细胞增多症 4 0

COVID-19 疫苗 阿斯利康 分析 打印
病例系列药物分析打印名称:COVID-19 疫苗阿斯利康分析打印报告运行日期:2021 年 12 月 2 日数据锁定日期:2021 年 12 月 1 日 18:30:04 MedDRA 版本:MedDRA 24.1 反应名称总计致命血液疾病贫血缺乏症贫血叶酸缺乏症 3 0 贫血维生素 B12 缺乏症 16 0 缺乏性贫血 1 0 缺铁性贫血 17 0 恶性贫血 6 0 贫血 NEC 贫血 177 0 自身免疫性贫血 1 0 失血性贫血 4 0 低色素性贫血 3 0 小细胞性贫血 1 0 正细胞性贫血 4 0 贫血溶血性 NEC 溶血性贫血 10 0 贫血溶血性免疫自身免疫性溶血性贫血 10 0 冷型溶血性贫血 1 0 埃文斯综合征 1 0 贫血 溶血性机械因素 非典型溶血性尿毒症综合征 1 0 红细胞碎裂综合征 1 0 出血倾向 瘀伤倾向增加 137 0 自发性血肿 8 0 自发性出血 2 0 凝血因子缺乏 获得性血友病 1 0 低纤维蛋白原血症 4 0 凝血病 凝血因子异常 12 0 抗磷脂综合征 29 0 凝血病 79 0 弥漫性血管内凝血 19 2 高凝状态 25 1 高纤维蛋白原血症 1 0 低凝状态 1 0 嗜酸性粒细胞疾病 嗜酸性粒细胞增多症 22 0 嗜酸性粒细胞增多综合征 1 0 血液囊肿和息肉 脾囊肿 1 0 血液疾病 血液疾病 14 1 骨髓疾病 2 0 骨髓水肿 1 0 肥大细胞活化综合征 10 0 肥大细胞增多症 3 0 高铁血红蛋白血症 1 0 溶血 NEC 溶血 11 0 白细胞增多症 NEC 白细胞增多症 4 0

COVID-19 阿斯利康疫苗分析 打印
病例系列药物分析打印名称:COVID-19 阿斯利康疫苗分析打印报告运行日期:2021 年 11 月 18 日数据锁定日期:2021 年 11 月 17 日 18:30:04 MedDRA 版本:MedDRA 24.1 反应名称总计致死血液疾病贫血缺乏症贫血叶酸缺乏症 2 0 贫血维生素 B12 缺乏症 15 0 缺乏性贫血 1 0 缺铁性贫血 17 0 恶性贫血 6 0 贫血 NEC 贫血 173 0 自身免疫性贫血 1 0 失血性贫血 4 0 低色素性贫血 3 0 小细胞性贫血 1 0 正细胞性贫血 4 0 贫血溶血性 NEC 溶血性贫血 10 0 贫血溶血性免疫性自身免疫性溶血性贫血 8 0 感冒型溶血性贫血 1 0 贫血 溶血性机械因素 非典型溶血性尿毒症综合征 1 0 红细胞碎裂综合征 1 0 出血倾向 瘀伤倾向增加 136 0 自发性血肿 8 0 自发性出血 2 0 凝血因子缺乏 获得性血友病 1 0 低纤维蛋白原血症 4 0 凝血病 凝血因子异常 11 0 抗磷脂综合征 29 0 凝血病 78 0 弥漫性血管内凝血 19 2 高凝状态 24 1 高纤维蛋白原血症 1 0 低凝状态 1 0 嗜酸性粒细胞疾病 嗜酸性粒细胞增多症 22 0 嗜酸性粒细胞增多综合征 1 0 血液囊肿和息肉 脾囊肿 1 0 血液病 血液疾病14 1 骨髓疾病 2 0 骨髓水肿 1 0 肥大细胞活化综合征 10 0 肥大细胞增多症 3 0 高铁血红蛋白血症 1 0 溶血 NEC 溶血 11 0 白细胞增多症 NEC 白细胞增多症 4 0 淋巴细胞增多症 11 0

COVID-19 阿斯利康疫苗分析 打印
病例系列药物分析打印名称:COVID-19 阿斯利康疫苗分析打印报告运行日期:2021 年 5 月 28 日数据锁定日期:2021 年 5 月 26 日 19:00:03 最早反应日期:1921 年 2 月 3 日 MedDRA 版本:MedDRA 24.0 反应名称总计致命血液疾病贫血缺乏症贫血维生素 B12 缺乏症 6 0 缺铁性贫血 6 0 恶性贫血 1 0 贫血 NEC 贫血 68 0 失血性贫血 2 0 小细胞性贫血 1 0 正细胞性贫血 1 0 溶血性贫血 NEC 溶血性贫血 8 0 溶血性免疫性贫血自身免疫性溶血性贫血 3 0 溶血性机械因素性贫血非典型溶血性尿毒症综合征 1 0 红细胞碎裂综合征 1 0 出血倾向 瘀伤倾向增加 84 0 自发性血肿 5 0 自发性出血 1 0 凝血因子缺乏 低纤维蛋白原血症 4 0 凝血病 凝血因子异常 7 0 抗磷脂综合征 22 0 凝血病 40 0 弥漫性血管内凝血 15 1 高凝状态 18 1 嗜酸性粒细胞疾病 嗜酸性粒细胞增多症 16 0 嗜酸性粒细胞增多症 1 0 血液系统疾病 血液疾病 6 0 骨髓疾病 2 0 骨髓水肿 1 0 肥大细胞活化综合征 5 0 肥大细胞增多症 1 0 高铁血红蛋白血症 1 0 溶血 NEC 溶血 8 0 白细胞增多症 NEC 白细胞增多症 3 0 淋巴细胞增多症 7 0 单核细胞增多症 2 0中性粒细胞增多症 13 0 白细胞减少症(NEC) 白细胞减少症 8 0 淋巴细胞减少症 8 0 淋巴系统疾病(NEC) 腹部淋巴结肿大 1 0 淋巴结纤维化 1 0 淋巴结疼痛 480 0 淋巴结炎 25 0

COVID-19 阿斯利康疫苗分析 打印
病例系列药物分析打印名称:COVID-19 阿斯利康疫苗分析打印报告运行日期:2021 年 6 月 3 日数据锁定日期:2021 年 6 月 2 日 18:30:03 最早反应日期:1921 年 2 月 3 日 MedDRA 版本:MedDRA 24.0 反应名称总计致命血液疾病贫血缺乏症贫血维生素 B12 缺乏症 7 0 缺铁性贫血 8 0 恶性贫血 1 0 贫血 NEC 贫血 79 0 失血性贫血 2 0 小细胞性贫血 1 0 正细胞性贫血 1 0 贫血溶血 NEC 溶血性贫血 8 0 贫血溶血免疫自身免疫性溶血性贫血 3 0 温型溶血性贫血 1 0 贫血溶血机械因素非典型溶血性尿毒症综合征 1 0 红细胞碎裂综合征 1 0 出血倾向 瘀伤倾向增加 89 0 自发性血肿 7 0 自发性出血 1 0 凝血因子缺乏 低纤维蛋白原血症 4 0 凝血病 凝血因子异常 8 0 抗磷脂综合征 23 0 凝血病 44 0 弥漫性血管内凝血 15 1 高凝状态 18 1 嗜酸性粒细胞疾病 嗜酸性粒细胞增多症 16 0 嗜酸性粒细胞增多症 1 0 血液囊肿和息肉 脾囊肿 1 0 血液病 血液病 7 0 骨髓疾病 2 0 骨髓水肿 1 0 肥大细胞活化综合征 5 0 肥大细胞增多症 1 0 高铁血红蛋白血症 1 0 溶血 NEC 溶血 8 0白细胞增多症 NEC 白细胞增多症 3 0 淋巴细胞增多症 8 0 单核细胞增多症 2 0 中性粒细胞增多症 13 0 白细胞减少症 NEC 白细胞减少症 9 0 淋巴细胞减少症 8 0 淋巴系统疾病 NEC 腹部淋巴结肿大 1 0
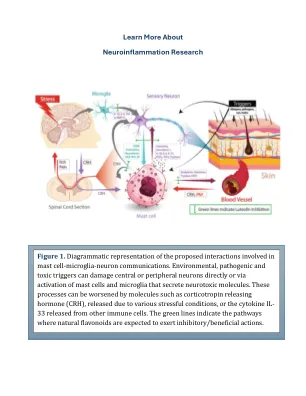
了解有关神经炎症研究的更多信息
,我们将使用Axion Maestro Edge Microectrode阵列(MEA)系统(Axion Biosystems,Atlanta,GA)刺激与内皮细胞,肥大细胞和小胶质细胞共同培养的HIPSC衍生神经元后的神经元健康(图。2)与试验键微流体系统(Netri,Lyon,France)结合起来,以创建人体类芯片模型。使用这种人体器官系统,我们将研究触发器的单个或组合的效果,以刺激由RNA-SEQ和酶联免疫吸附测定(ELISA)分析的神经毒性分子的释放(ELISA),该分子(ELISA)专注于炎症,神经元连通性和血管分布(图。2)。由IPSC组成的人类器官已用于研究某些神经系统疾病,也可以用于筛查潜在治疗。