XiaoMi-AI文件搜索系统
World File Search System膜电位
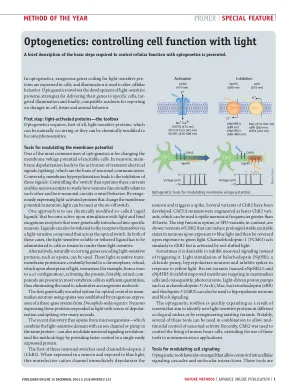
光遗传学:用光控制细胞功能
调节膜电位的工具 光遗传学最常见的用途之一是改变可兴奋细胞的膜电位。在神经元中,膜去极化会导致瞬态电信号(脉冲)的激活,这是神经元通讯的基础。相反,膜超极化会导致这些信号的抑制。控制操作这些电流的“开关”使神经科学家能够研究神经元在功能上如何相互关联以及神经元回路如何控制行为。通过外源表达改变神经元膜电位的光激活蛋白,光可以用作开关。一种方法是使用化学修饰的所谓“笼状配体”,这些配体在光刺激下变得活跃并与通过基因引入特定神经元的外源性受体结合。配体也可以通过充当光开关的光敏化合物与受体本身相连。在这两种情况下,都必须将光敏的可溶性或束缚配体注入细胞或组织,使它们对光敏感。或者,可以使用编码光敏蛋白(如视蛋白)的天然基因。这些光敏跨膜蛋白与发色团视网膜共价结合,视网膜吸收光后发生异构化(例如,从反式变为顺式构型),从而激活蛋白质。值得注意的是,视网膜化合物在大多数脊椎动物细胞中含量充足,因此无需注入外源分子。第一个利用视蛋白进行哺乳动物神经元光学控制的遗传编码系统是通过外源表达果蝇的三基因系统建立的。表达这些蛋白质的神经元对光的反应是数秒内的去极化和尖峰波。最近发现,微生物中的视蛋白(将光敏域与同一蛋白质中的离子通道或泵相结合)也可以调节神经元信号,通过在单个易于表达的蛋白质中提供更快的控制,彻底改变了该方法。这些神经元开关中的第一个使用了通道视紫红质-2 (ChR2)。当在神经元中表达并暴露于蓝光时,这种非选择性阳离子通道会立即使神经元去极化

意识的先验基础
考虑以下场景:你是一位身无分文的哲学家,遇到了一位奇怪的超人类主义脑科学家。他愿意为你提供一大笔金钱奖励,让你参与他的实验,该实验使用了一种新型的脑部扫描仪。同意参与后,你签署了一份声明,允许他拥有从扫描中获得的所有信息。他让你进入一个特殊的房间,而他则在另一个房间等候。你注意到你所在的房间只有一扇红门和一扇蓝门。然后你接到了超人类主义者的视频电话。他向你解释说扫描已成功完成,他刚刚使用了有关你脑细胞的所有详细信息来创建一个相当精确的大脑模拟,该模拟目前正在一台强大的超级计算机上实时运行。这个模拟器是一个复杂的软件,可以模拟你脑细胞的精确虚拟 3-D 模型的膜电位。不仅是你的大脑,事实上你的整个身体和房间都在用它模拟。由于你不相信他,他重新连接了视频通话,这样你就可以和模拟的自己交谈了。然后,你与这个模拟的自己交谈了很长时间,直到你确定这个模拟确实在功能上与你相同,并且拥有与你相同的记忆。模拟的自己相信自己是真实的,并相信你是模拟的。你们不同意,并就这个问题进行了长时间的争论。最后,你们俩都不得不承认,目前你们俩都不确定谁是原版,谁是复制品。在你了解到这一点后,超人类主义者重新加入视频通话,并解释说,承诺的金钱奖励可以通过离开红门获得;但如果你从蓝门离开,你就会身无分文。当你开始走向红门时,他继续解释说,模拟的设置是这样的,当你从蓝门离开时,模拟的你将被无痛地、甚至没有注意到地终结。然而,如果它从红门离开,它将遭遇痛苦的死亡。由于你并不真正知道你是否被模拟了,你从那扇红门退了回来。

心肌缺血和再灌注损伤的病理生物学
摘要:本评论提出了一种综合方法,用于分析心肌缺血和再灌注损伤,以及在急性心肌梗死(AMI)和其他临床环境演变过程中心肌条件的调节影响。实验研究涉及一系列体外,体内和体内模型,并且已经为进行严格的临床前研究制定了指南,并确定了各种形式的细胞损伤和死亡,而在不断发展的AMI中。ami体内由肿瘤(细胞损伤肿胀)主导,导致尸检和缺血性心肌细胞(CMC)的最终坏死,而没有收缩带形成或没有收缩带形成。冠状动脉闭塞后,再灌注剂量大量的脑膜内心肌心肌症,而再灌注损伤则占最终心脏心脏心脏梗死的50%。AMI进展是由损伤(或危险)相关的分子模式(也称为Alarmins)介导的,该分子模式也称为Alarmins,它激活了模式识别受体并启动炎症反应。在临床前研究中,由于对CMC和微脉管系统的影响,可以通过药理学或物理手段进行预处理或后的后期来预防致命的再灌注损伤。 调节涉及触发因素,胞质介质和细胞内效应子。 线粒体在CMC的可行性维持和丧失中具有核心作用。 严重渗透心肌的再灌注会导致线粒体渗透能力过渡孔(MPTP)的持续开放。 调节会阻止MPTP的持续开放。在临床前研究中,由于对CMC和微脉管系统的影响,可以通过药理学或物理手段进行预处理或后的后期来预防致命的再灌注损伤。调节涉及触发因素,胞质介质和细胞内效应子。线粒体在CMC的可行性维持和丧失中具有核心作用。严重渗透心肌的再灌注会导致线粒体渗透能力过渡孔(MPTP)的持续开放。调节会阻止MPTP的持续开放。打开MPTP后,线粒体膜电位(ΔψM)迅速丢失,能量产生停止。将策略转换为患者的临床管理一直在努力。在开放心脏手术和心脏移植期间,讨论了将实验发现转换为调节和改善缺血和改善方法的方法的状态。

使用脑电图 (EEG) 信号预测癫痫发作的机器学习方法
活动:a) 神经元膜的快速去极化,这取决于钠离子和钾离子的电压,并产生动作电位 (AP) [3]。b) 由于突触活动和几种神经递质系统的功能,膜电位变化较慢 [4]。AP 是膜电位的快速变化,时间为 1 – 2 毫秒,它使细胞内电位从负变为正,并迅速返回细胞内静息电位。它具有较小的场电位分布(在细胞外环境中的渗透较少)并且持续时间较短(约 1 毫秒,而突触后电位则为 15 到 200 多毫秒)[3,5]。突触后电位扩散到颅骨表面并且可以测量。为了测量大脑活动,我们可以使用一种非侵入性的方法,通过一种称为脑电图 (EEG) 的设备将电极放在头皮上。EEG 信号主要由突触后锥体细胞的可测量电位产生,这些细胞彼此平行且垂直于颅骨表面。它创建了一个细胞外皮质偶极层 [5,6]。因此,颅骨上的电极代表皮质神经元突触后电位的时间和位置。它还包括大皮质区域中缓慢且同时的电位变化(图 1)[7,8]。EEG 信号可用于识别许多临床问题,例如精神分裂症、阿尔茨海默病、失眠症、睡眠障碍、癫痫症、脑瘤和中枢神经系统感染。除了具有非侵入性和精确的时间分辨率之外,该技术还具有低成本且不需要极端的安全限制 [5]。通过脑电图信号可以发现,癫痫发作通常是自发性的。它们是由部分脑细胞突然放电引起的,因此会导致大脑暂时兴奋。有时癫痫发作可能会被忽视,或者可能与其他脑部疾病(如脑膜炎或中风)混淆,这些疾病也会导致相同的症状。研究表明,大约 10% 的人一生中至少会经历一次癫痫发作 [ 10 ]。对脑电图 (EEG) 信号中的癫痫进行精确分析可以揭示有关这种普遍存在的脑部疾病的宝贵事实 [ 11 ]。由于 EEG 信号非常复杂,因此需要分析多种因素。手动目视检查 EEG 信号已被发现有助于识别模式。然而,这种方法需要高水平的技术和分析能力,以及多种信号处理技术[12]。因此,近年来,癫痫发作的自动检测

下一代神经质量模型
摘要 在本论文中,我们介绍了下一代神经质量模型的新颖扩展和应用。 Montbrió、Pazó 和 Roxin (MPR) 已证明,二次积分和放电 (QIF) 神经元集合的集体行为可以用平均膜电位和放电率来精确描述,从而将无限大的微观网络的问题维度降低为低维宏观描述。由于神经质量提供了平均膜电位的途径,因此它可以作为局部场电位和脑电图信号的指标。本论文的贡献之一是在 MPR 模型中实现短期突触可塑性(STP)。基于工作记忆 (WM) 的突触理论,我们在多群体设置中使用 QIF 网络及其精确的平均场边界重现了 WM 的机制。实验中观察到,神经质量模型在记忆加载和维持过程中表现出 β-γ 带的振荡,而我们在启发式模型中遇到空的 β-γ 带。此外,我们指出了这些功率带是如何由基频之间的共振形成的,并与记忆中保留的元素数量相关。我们还对大约五种元素的最大 WM 容量进行了分析估计。第二个贡献是应用多种群模型来检验癫痫发作传播的临床假设。我们使用从健康受试者和癫痫患者的扩散 MRI 扫描获得的结构连接组。我们描述了如何将类似癫痫发作的事件建模为从低活动状态到高活动状态的募集。外部输入可以触发此类事件并导致一系列招募,从而模仿危机的时空传播。数值结果表明,癫痫患者对延长招募事件比健康受试者更敏感。我们还发现,我们的模型中首先招募的大脑区域与招募的次级网络的手术前评估之间存在良好的一致性。作为第三个贡献,我们使用慢-快动力学研究了 STP 存在下的神经网络和质量。根据施加到群体的慢周期电流的幅度,集体行为可以处于亚阈值振荡状态,也可以处于爆发状态,即在准静态漂移和大幅度快速振荡之间交替。这两个区域之间有一个狭窄的参数间隔,就像鸭子爆炸一样。在这个区域,我们报告了跳跃式鸭翼,它接近通常排斥的不变集。对于中间时间尺度分离,爆发通过混合型环面鸭翼组织的尖峰添加机制以连续的方式出现,其轨迹接近排斥平衡和极限环家族。为了实现更强的时间尺度分离,连续过渡被跳跃式鸭翼阻挡。在神经团中观察到的机制也是导致网络爆发的原因。总而言之,本论文将下一代神经质量模型置于神经科学建模的更广泛背景中,并为未来的工作提供了新的视角。这包括考虑以下方法

金属和细胞氧化还原和免疫状态
我们非常高兴地在“稳态:金属和蜂窝氧化还原和免疫状态”上介绍了这个特刊。这个问题的目的是探索金属稳态,细胞氧化还原平衡和免疫功能之间的复杂关系。保持适当的金属稳态和细胞氧化还原平衡的重要性不能被夸大。金属在许多生物过程中起着关键作用,包括酶促反应,信号转导和DNA复制。但是,金属浓度的失衡会导致细胞损伤和功能障碍。同样,氧化剂和抗氧化剂的平衡对于细胞健康至关重要。太多的氧化剂会导致氧气应激,而过量的抗氧化剂会破坏信号通路。重要的是,金属稳态和细胞氧化还原平衡都与免疫功能紧密相关。金属离子在免疫细胞信号传导和分化中起重要作用,而细胞氧化还原平衡会影响免疫细胞的激活和增殖。金属家居和细胞氧化还原平衡的破坏会导致免疫功能受损,并增加对感染的敏感性。此收集中的第五篇文章报告了严重疾病发病机理期间干涉稳态的分子机制。第一篇文章“突触活动通过铁代谢来增强神经元生物能力”,Tena Morraja等人。[1]表明,突触活性会触发铁代谢基因的转录上调,从而导致细胞和线粒体铁的摄取增强。铁可用性的这种增加为电子传输链配合物提供了促进,从而促进了线粒体生物能学的长期改善。实际上,当抑制线粒体铁转运蛋白MFRN1时,活性介导的生物能力的增强被阻断。为了更好地理解突触活动对神经元代谢的持久影响,他们探索了刺激神经元中线粒体能量学的变化。结果表明,线粒体膜电位和消耗量增加,MFRN1的表达受到CREB的调节,Creb是突触可塑性的关键调节剂。这表明突触可塑性程序的表达与满足能量需求相关的增加所需的表达。Michaelis等人的第二个手稿是“胎盘锰和铁转移的差异和相互作用”。[2]研究了锰(Mn)和铁(Fe)在Bewo B30滋养细胞层中的转移。这些元素在胎儿发育中起着至关重要的作用,但是宫内过多的MN与不良妊娠结局有关。这项研究揭示了MN和Fe的胎盘转移有着明显的差异,MN转移在很大程度上独立于应用剂量。同时暴露两个元素表明它们具有共同的转移机制。作者认为,MN的转移可能涉及主动和被动传输过程的组合,因为尽管暴露了不同的情况,但在BOWO细胞中DMT1,TFR或FPN仅略有改变。Reinert等人的第三篇文章。铁是能量代谢中的关键元素,但是当Fe 2+ /Fe 3+比率出现问题时,它可能会产生不利影响。[3]探索安全的铁处理。