XiaoMi-AI文件搜索系统
World File Search System蛋白



Rho GTPases及其效应蛋白
使用的缩写:ACK,激活的CDC42相关酪氨酸激酶; GEF,鸟苷核苷酸交换因子; PH,Pleckstrin同源性; DH,DBL同源性; PIP 2,磷脂酰肌醇4,5-双磷酸;间隙,GTPase激活蛋白; GDI,鸟苷核苷酸解离抑制剂; SRF,血清反应因子; NF-κB,核因子κB; Jnk,c-jun n末端激酶;婴儿床,cdc42/rac-Interactive结合; REM,Rho ectector同源性; RKH,ROK – Kinectin同源性; MLC,肌球蛋白轻链; PI-4-P5K,磷脂酰肌醇-4-磷酸5-激酶; GTP [s],鸟嘌呤5« - [γ -thio]三磷酸; MAP激酶,有丝分裂原激活的蛋白激酶; MLK,混合细胞激酶; ACC,反平行线圈; BTK,布鲁顿的酪氨酸激酶; MBS,肌球蛋白结合亚基; ERM,Ezrin/radixin/Moesin; FH,形态学;黄蜂,Wiskott-Aldrich-Syndrome蛋白;波浪,黄蜂样的垂直蛋白质蛋白; lim激酶; EGF,表皮生长因子; TNFα,肿瘤坏死因子α; Mekk,地图激酶激酶激酶; PAK,P21激活的激酶; PKN,蛋白激酶N; MRCK,肌发育症激酶相关的CDC42结合激酶。1应向谁致辞(电子邮件Anne.bishop!ucl.ac.uk)。

基于丝素蛋白的柔性电子产品...
随着物联网 (IoT) 的快速发展和 5G 的引入,传统的硅基电子产品已无法完全满足市场需求,例如由于机械不匹配导致的非平面应用环境。这为使用柔性材料避免物理刚性的柔性电子产品带来了前所未有的可能性。丝素蛋白、纤维素、果胶、壳聚糖和黑色素因其出色的生物相容性和生物降解性而成为下一代柔性电子产品最有吸引力的材料之一。丝素蛋白在生物相容性和生物降解性方面优于它们,并且还具有多种其他理想特性,例如可调节的水溶性、出色的光学透射率、高机械弹性、重量轻和易于加工,而这些特性是其他材料部分或完全不具备的。因此,丝素蛋白已成为生物相容性柔性电子产品最广泛使用的构建块之一,尤其是用于可穿戴和可植入设备。此外,近年来,丝素蛋白的功能特性研究也越来越受到重视,如介电特性、压电特性、高失电子倾向性、环境敏感性等。本文不仅介绍了不同种类丝素蛋白的制备技术以及丝素蛋白作为基础材料应用的最新进展,还介绍了丝素蛋白作为功能元件的最新进展。本文还对丝素蛋白基柔性电子产品面临的挑战和未来发展进行了探讨。

核篮蛋白调节1
核孔复合物(NPC)介导细胞核和细胞质之间的所有流量,是细胞中最稳定的蛋白质组件之一。有趣的是,发芽的酵母菌细胞具有两个NPC的两个变种,它们在存在或不存在核篮蛋白MLP1,MLP2和12 PML39的情况下有所不同。这些篮子蛋白的结合发生在NPC组装中很晚,而MLP阳性NPCS 13被排除在与核仁接壤的核包膜区域中。14在这里,我们使用重组诱导的TAG交换(RITE)来研究单个NPC中所有NPC 15子复合物的稳定性。我们表明,核篮蛋白MLP1,MLP2和16 PML39通过多个细胞分割循环与NPC保持稳定,并且MLP1/2是17负责将NPC从核方区域排除。此外,我们证明了NUP2的18结合还通过独立途径从该区域耗尽了MLP阴性NPC。我们19开发了一种在萌芽酵母中进行单个NPC跟踪的方法,并观察到在没有核篮成分的情况下,NPC在没有核篮成分的情况下表现出20个迁移率。我们的数据表明,NPCS 21在核上的分布受核篮蛋白与核内部的相互作用的控制。22

融合蛋白的人工智能三维...
融合蛋白由至少两个部分蛋白结构域的组合组成,每个蛋白结构域由单独的基因编码,合并以进行关节转录和转移。精确预测蛋白质的三维(3D)结构对于随后的药物发现过程的准确性至关重要。这包括预测蛋白质功能,研究蛋白质 - 蛋白质相互作用,查找抑制剂,设计抗体和分析蛋白质 - 配体相互作用。1,2蛋白质3D结构的预测主要依赖于蛋白质序列数据及其结构同源性。3巨大的努力以更好地理解并研究野生型(WT)蛋白的3D结构。但是,我们缺乏预测融合蛋白3D结构并具有足够知识的努力。融合蛋白。此过程是由DNA双链断裂引起的染色体重排触发的。这些融合蛋白是两种不同蛋白质结构(包括主要功能结构域或部分)的球状形式的组合形式,可能会导致细胞中具有新功能或调节的新型蛋白质,从而导致疾病。4许多融合蛋白已被用作癌症治疗中的治疗靶标。5–8
分子口服GLP-1蛋白
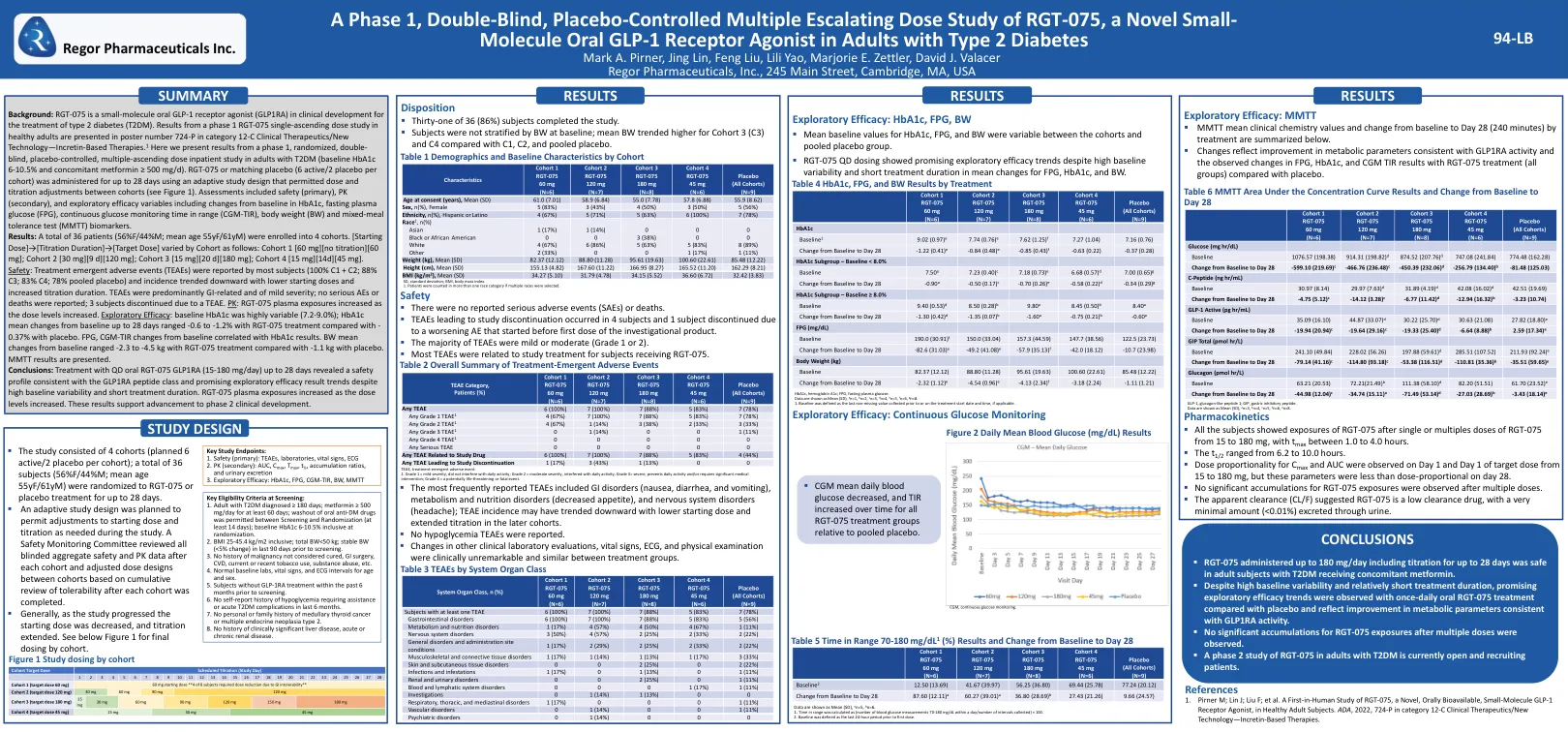
背景:RGT-075是一种用于治疗2型糖尿病(T2DM)的临床发育中的小分子GLP-1受体激动剂(GLP1RA)。从健康成年人中的1阶段RGT-075单级剂量研究的结果是,在12-C类临床治疗剂/新技术(基于Incretin的疗法)中,海报编号为724-P。1此处,我们介绍了1阶段的结果,即随机,双盲,安慰剂对照的,多重固定剂量的住院治疗研究(基线HBA1C 6-10.5%),伴随二甲双胍≥500mg/d)。RGT-075或匹配的安慰剂(每个队列的6个活性/2个安慰剂)使用自适应研究设计最多28天,该设计允许剂量和滴定调整,并调整同类群之间(见图1)。评估包括安全性(主要),PK(次要)和探索性功效变量,包括HBA1C中基线的变化,禁食血浆葡萄糖(FPG),范围内连续的葡萄糖监测时间(CGM-TIR),体重(BW)(BW)和混合耐受性测试(MMTT)生物剂。结果:总共36例患者(56%f/44%m;平均年龄为55yf/61ym)被招募为4个队列。[起始剂量]→[滴定持续时间]→[靶剂量]通过队列变化如下:队列1 [60 mg] [无滴定] [60 mg];队列2 [30 mg] [9 d] [120 mg];队列3 [15 mg] [20 d] [180 mg];队列4 [15 mg] [14d] [45 mg]。安全:大多数受试者报告了治疗出现的不良事件(TEAE)(100%C1 + C2; 88%C3; 83%C4; 83%C4; 78%合并安慰剂),并且发病率下降,起始剂量较低,滴定持续时间增加。PK:随着剂量水平的增加,RGT-075血浆暴露量增加。MMTT结果提出。茶叶主要与胃肠道相关,严重程度轻度;没有严重的AE或死亡报告; 3个受试者因茶而停产。探索功效:基线HBA1C变化很大(7.2-9.0%); HBA1C的平均基线变化从基线到28天的变化范围为-0.6至-1.2%,而RGT -075治疗的变化为-0.37%,而安慰剂为-0.37%。FPG,CGM-TIR与基线的变化与HBA1C结果相关。bw的平均基线变化范围为-2.3范围为-4.5 kg,而RGT -075治疗的平均变化为-1.1 kg,安慰剂为-1.1 kg。结论:用QD口服RGT-075 GLP1RA(15-180 mg/天)的处理长达28天,显示出与GLP1RA肽类别的安全性,尽管基线变异性很高,但仍具有GLP1RA肽类和有希望的探索性结果趋势。随着剂量水平的增加, RGT-075血浆暴露量增加。 这些结果支持第2阶段临床发展的进步。RGT-075血浆暴露量增加。这些结果支持第2阶段临床发展的进步。

病毒衣壳蛋白的免疫原性
病毒衣壳蛋白被广泛用于亚单位疫苗开发,但其生产复杂性和低免疫原性常常阻碍其发展。在这里,我们报告了一种通过将 mRNA 疫苗技术与蛋白质工程相结合来克服这些挑战的简单方法。以非洲猪瘟病毒 (ASFV) 衣壳蛋白 P72 和五邻体为模型,我们将它们设计成膜结合和分泌形式,并通过 mRNA 疫苗接种将它们的免疫原性与小鼠和猪的天然细胞内形式进行了比较。膜结合和分泌的 P72 和五邻体独立于病毒伴侣折叠成其天然多聚结构,因此保留了它们的构象表位。膜结合的 P72 和五邻体也比其分泌或细胞内的对应物引起明显更强的抗体和 T 细胞反应。我们的研究提供了一种简单的方法 27 来增强病毒衣壳蛋白的折叠、多聚体结构形成和免疫原性 28,用于 ASFV 亚单位疫苗的开发和一般细胞内蛋白质的免疫原性。29 30 31
