XiaoMi-AI文件搜索系统
World File Search System诱导

诱导多能干细胞瘤变的研究进展
TI方向分化潜力(ESC),并避免了ESC的伦理问题。自IPSC发明以来,它已迅速应用于疾病建模,药物开发,再生医学和基因调节中,尤其是在再生医学研究领域。但是,IPSC移植后肿瘤已成为使用IPSC进行再生医学的主要障碍,因此IPSC中的肿瘤已成为当前IPSC研究中的热门问题。本文简要审查了IPSC和肿瘤细胞之间的关系,移植后IPSC的恶性转化以及如何减少其以及IPSC的体内监测技术。

激光诱导制备芳纶纤维基石墨烯的探索及研究
Technology, 2021, 201: 108541.[19] Steinke K, Groo L, Sodano H A. Laser induced graphene for in situ ballistic impact damage and delamination detection in aramid fiber reinforced composites [J].Composites Science and Technology, 2021, 202: 108551.[20] 杜晓云 , 李金宝 , 杨斌 , 等 .芳纶树脂液浸渍协同冷压 光制备高强度间位芳纶纸的研究 [J].中国造纸 , 2024, 43(4): 120 - 129.Du X Y, Li J B, Yang B, et al.Study on preparing high strength meta - aramid paper by aramid resin solution impregnation combined with cold pressing[J].China Pulp & Paper, 2024, 43(4): 120 - 129.[21] 关振虹 , 李丹 , 宋金苓 , 等 .易染间位芳纶的制备及其 性能 [J].纺织学报 , 2023, 44(6): 28 - 32.Guan Z H, Li D, Song J L, et al.Preparation and properties of dyeable meta - aramid fiber[J].Journal of Textile Research, 2023, 44(6): 28 - 32.[22] 朱文豪 , 宋欢 , 丁娉 , 等 .沉析纤维长度对间位芳纶纸 性能的影响 [J].中国造纸 , 2024, 43(1): 109 - 115.

通过诱导seq
图2:与替代性DSB测序技术相比,诱导seq表现出无与伦比的灵敏度和动态范围。(a)诱导seq同时检测高度复发的诱导DSB和低级内源性DSB,并以高分辨率。基因组浏览器视图(IgV)诱导seq读取映射到HEK293T细胞的10MB部分,随后与限制性核酸内核酸内切酶Hindiii进行原位裂解。(顶部面板)高度复发性酶诱导的断裂表示在低分辨率(10MB,0-1000读物)时的绝大多数读数。(底部面板)高分辨率视图(粉红色的亮点,500kb,0-20读取)显示出未处理样品中存在的低水平的单源性断裂,以及在复发性印度诱导的突破(绿色亮点)中。(b)诱导seq读取的映射在Hindiii目标位点显示了断裂两侧的单核苷酸断裂映射的精度。(c)对经过治疗和对照样品的每个细胞测量的断裂定量。诱导seq在样品之间的3个数量级上定量检测到每个细胞的断裂。(d和e)通过酶Hindiii和ecorv检测体外裂解限制位点时诱导seq和dsbapture之间的比较。(d)使用诱导seq映射到测序和对齐基因组的读取和对齐基因组的比例更大。(e)使用少800倍的细胞,诱导seq鉴定了与DSBCAPTURE确定的ECORV(93.7%)相似的Hindiii限制位点(92.7%)。(f)使用诱导seq的诱导DSB检测的动态范围。除了在印度内目标序列(AAGCTT)上鉴定出的断裂外,还确定了多个1BP和2BP不匹配靶向位点。诱导seq测得的诱导的休息事件,跨越了8个数量级,从在印度内靶标地点确定的约1.5亿次断裂到最少频繁的脱离目标的5个断裂。(g)在检测ASISI诱导的活细胞中诱导的疾病,DSBCAPTURE和BLISS之间的比较。将测序的读取数(顶部面板)与每个实验(底部面板)识别的ASISI位点的数量进行了比较。诱导seq使用比DSBCAPTUE少的40倍读数检测到最大数量的ASISI位点,而读取的读数比Bliss少23倍。

通过丙酸糖醇结合诱导铁凋亡诱导...
丙酸丙酸酯(CP)最初由美国食品药品监督管理局(FDA)批准,用于治疗由于其抗炎症特性而导致的湿疹和牛皮癣等皮肤状况,已成为在Keap-1中以突变为特征的肺癌中的肺癌症的有前途的候选者,在Keap-1中,负责为n ragencultator n n nrf-2 [2] [2] [2] [2]。NRF-2的上调与肺癌患者的预后不良有关,影响了大约三分之一的非小细胞肺癌(NSCLC)。此外,暴露于辐射还激活了NRF-2导致放射线[3,4]。针对NRF-2的小分子抑制剂在使癌细胞对化学疗法的敏感性方面表现出了希望,这表明它们作为放射疗法的佐剂潜力[5]。因此,在当前研究中,CP与辐射相结合,以评估其对Keap-1突变体肺癌细胞敏感的潜力。用CP抑制NRF-2并暴露于辐射促进的铁凋亡诱导,从而增强了NSCLC细胞的放射敏性[6]。铁凋亡,一种由铁内脂质过氧化物诱发的非凋亡细胞死亡的铁依赖性形式,是

886-quizartinib-daunorubicin and-cytarabine诱导 -
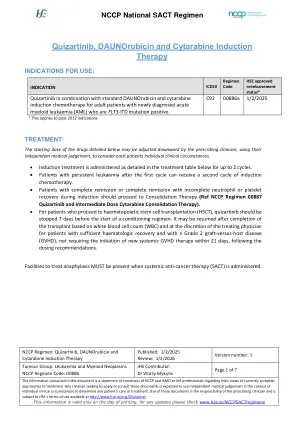
诱导治疗是在下面的处理表中详细介绍的,最多2个周期。第一个周期后患有持续性白血病的患者可以接受第二个诱导化疗周期。在诱导过程中,患有完全缓解或完全缓解的患者应进行巩固治疗(参考NCCP方案00887 Quizartinib和中等剂量的细胞蛋白滨固结疗法)。对于继续进行造血干细胞移植(HSCT)的患者,应在调节方案开始前7天停止Quizartinib。 可以根据白细胞计数(WBC)完成移植后恢复,并由治疗医师自行决定,患有足够的血液恢复并患有≤2级移植物 - 主疾病(GVHD)的患者,不需要在21天内进行新的全身GVHD治疗,并不需要在21天内进行启动。对于继续进行造血干细胞移植(HSCT)的患者,应在调节方案开始前7天停止Quizartinib。可以根据白细胞计数(WBC)完成移植后恢复,并由治疗医师自行决定,患有足够的血液恢复并患有≤2级移植物 - 主疾病(GVHD)的患者,不需要在21天内进行新的全身GVHD治疗,并不需要在21天内进行启动。

诱导血管性水肿
血管水肿每年在美国(美国)进行100,000多个急诊科(ED)访问,约有11%的患者需要入院[1]。这种短暂的,非固定的水肿可能会影响各种地点,并且可能会发展为威胁生命的气道水肿,如果不迅速治疗,需要插管。在大多数患者中,血管性水肿的原因尚不清楚。但是,病理生理学可以归因于组胺或心动激肽介导的机制。组胺介导的血管性水肿是响应于I型免疫球蛋白E(IgE)超敏反应的肥大细胞脱粒的结果,占所有病例的40%至70%。虽然缓激肽介导的血管性水肿不太常见,但由于上呼吸道的显着参与,这些病例可能持续且更严重。Bradyinin介导的血管性水肿可以进一步分类为遗传性血管性水肿(HAE),获得性血管性水肿或血管紧张素转化酶(ACE)抑制剂诱导的血管性水肿(ACEI-AE)[2]。

文章通过诱导ACT
肿瘤微环境的免疫抑制是有助于肿瘤进展和免疫疗法抗性的关键因素。启动肿瘤免疫微环境(时间)已成为改善癌症免疫疗法效率的有前途的策略。在这项研究中,我们研究了非毒性射频辐射(RFR)暴露对肿瘤进展和时间表型的影响,以及在肺转移性黑色素瘤模型(PMM)模型中PD-1阻滞的抗肿瘤潜力。PMM的小鼠模型是通过尾静脉注射B16F10细胞建立的。 从注射后的第3天开始,将小鼠以平均特定的吸收率为9.7 W/kg,每天1小时,持续14天。 RFR暴露后,收集肺组织,并提取RNA进行转录组测序。分离PMM - 纤维化免疫细胞进行单细胞RNA-seq分析。 我们表明,RFR暴露显着阻碍了PMM进展,并通过改变肿瘤 - 纤维编织免疫细胞的比例和转录表现,并伴随着PMM的重塑时间。 rfr暴露增加了肿瘤 - 纤维化CD8 + T细胞的激活和细胞毒性特征,尤其是在早期激活子集中,具有与T细胞细胞毒性相关的上调基因。 CD8 + T细胞中RFR暴露在PD-1检查点途径上调。 RFR暴露还增加了NK细胞亚群,并在PMM中具有增加的细胞毒性特征。 rfr暴露增强了肿瘤 - 纤维纤维CD8 + T细胞和NK细胞的效应子功能,从而证明了细胞毒性分子的表达增加。PMM的小鼠模型是通过尾静脉注射B16F10细胞建立的。从注射后的第3天开始,将小鼠以平均特定的吸收率为9.7 W/kg,每天1小时,持续14天。RFR暴露后,收集肺组织,并提取RNA进行转录组测序。分离PMM - 纤维化免疫细胞进行单细胞RNA-seq分析。我们表明,RFR暴露显着阻碍了PMM进展,并通过改变肿瘤 - 纤维编织免疫细胞的比例和转录表现,并伴随着PMM的重塑时间。rfr暴露增加了肿瘤 - 纤维化CD8 + T细胞的激活和细胞毒性特征,尤其是在早期激活子集中,具有与T细胞细胞毒性相关的上调基因。CD8 + T细胞中RFR暴露在PD-1检查点途径上调。RFR暴露还增加了NK细胞亚群,并在PMM中具有增加的细胞毒性特征。rfr暴露增强了肿瘤 - 纤维纤维CD8 + T细胞和NK细胞的效应子功能,从而证明了细胞毒性分子的表达增加。RFR激活的CD8 + T细胞和NK细胞介导 RFR诱导的PMM生长抑制作用。 我们得出的结论是,非侵入性RFR暴露会诱导时间的抗肿瘤重塑,从而导致抑制肿瘤进展,这为时间启动和潜在的与癌症免疫疗法结合提供了有希望的新型策略。RFR诱导的PMM生长抑制作用。我们得出的结论是,非侵入性RFR暴露会诱导时间的抗肿瘤重塑,从而导致抑制肿瘤进展,这为时间启动和潜在的与癌症免疫疗法结合提供了有希望的新型策略。

报告 - 用于诱导多...
摘要:澳大利亚有明确的愿望成为化石燃料的替代氢的主要出口国,也是减少CO 2排放的一部分,如联邦和州政府于2019年共同发布的国家氢策略所规定的那样。在2021年,澳大利亚能源市场运营商指定了第一次名为“氢超级大国”的网格预测场景。澳大利亚不仅希望通过建立一个新的出口行业来利用日本和韩国等地方对零碳氢的需求,而且还需要减轻其从煤炭和液化天然气(例如日本)等主要客户出口收入的内置碳风险,例如日本和韩国,以使其能源系统脱氧。这将氢置于能源,气候变化和经济增长的联系,对能源安全的影响。关于该主题的许多已发表文献都集中在主要的氢出口商外观以及将需要采取哪些步骤来实现它的细节上。但是,在能源安全和出口经济脆弱性方面,研究对澳大利亚国内能源体系的影响似乎存在差距。本文的目的是为成为澳大利亚能源系统的主要氢出口商的含义开发一个概念框架。比较了澳大利亚的各种绿色氢出口方案,最新和全面的选择是进一步检查动物能源系统影响的基础。在这种情况下,估计需要248.5 GW的新可再生电力发电能力到2050年需要产生2088 PJ绿色氢氢的ElectrolySer输出所需的额外的867 TWH,以占该时间的55.9%的澳大利亚总电力需求。比较出口资源的特征及其与国内经济和能源系统的相互作用。对这些现有的资源出口框架进行了审查,以适用于特定因素对出口为导向的绿色氢生产的适用性,然后将适用的因素汇编成一个新颖的概念框架,以从大规模出口的绿色氢出口中出口国内影响。然后,使用既定的能源出口商脆弱性和国内能源安全的指标对绿色氢出口超级大国(2050)方案进行定量评估,并将其与澳大利亚2019年的2019年能源出口投资进行了比较。这项评估发现,在几乎所有因素中,出口商脆弱性都会减少,并且通过从化石燃料出口到绿色氢的过渡增强了国内能源安全,除了国内能源系统暴露于国际市场力量外。
