XiaoMi-AI文件搜索系统
World File Search System负反馈


生长介导的负反馈形状定量抗生素反应
剂量 - 响应关系是从分子到整个细胞水平的多个尺度上描述生物系统的一般概念。临床上相关的例子是对抗生素的细菌生长反应,该反应通常以剂量 - 反应曲线为特征。剂量 - 反应曲线的形状在抗生素之间差异很大,并且在治疗,药物相互作用和耐药性演化中起关键作用。但是,构成剂量的机制 - 反应曲线在很大程度上不清楚。在这里,我们在大肠杆菌中表明,明显的浅剂量 - 甲甲化抗生素的响应曲线是由负生长介导的反馈回路引起的:甲氧苄啶减慢了生长,从而削弱了这种抗生素的作用。在分子水平上,这种反馈是由药物靶标的二氢叶酸还原酶(Fola/dhfr)的上调引起的。我们表明,这种上调不是对甲氧苄啶的特定响应,而是遵循的普遍趋势线,主要取决于增长率,而与其原因无关。重新布线反馈回路以可预测的方式改变了剂量 - 响应曲线,我们使用细胞资源分配和生长的数学模型来证实这一点。我们的结果表明,生长介导的反馈回路可能会更普遍地塑造药物反应,并且可以利用设计为设计的陷阱,从而可以选择抵抗药物耐药性。

长链非编码 RNA LUCAT1 是人类干扰素反应的负反馈调节剂
长链非编码 RNA 是包括免疫反应在内的生物过程的重要调节因子。lncRNA 的免疫调节功能主要在小鼠模型中得到揭示,而对 lncRNA 在人类免疫反应中的了解有限。在这里,我们鉴定出 lncRNA LUCAT1,它在受脂多糖和其他先天免疫刺激刺激的人类髓系细胞中上调。在髓系细胞中靶向删除 LUCAT1 会增加响应 LPS 的 I 型干扰素刺激基因的表达。相反,增加 LUCAT1 表达会导致可诱导的 ISG 反应降低。在活化细胞中,LUCAT1 在细胞核中富集,并与染色质结合。此外,LUCAT1 通过与细胞核中的 STAT1 相互作用来限制干扰素刺激基因的转录。总之,我们的研究强调了 lncRNA LUCAT1 作为诱导后反馈调节因子的作用,其功能是抑制人类细胞的免疫反应。

miR-21 与 PPAR- 在透明肾细胞癌中的双重负反馈相互作用
1 里尔大学,法国国立科学研究院,法国国家健康与医学研究院,里尔临床医学院,UMR9020-U1277—CANTHER—癌症异质性可塑性和治疗耐药性,F-59000 里尔,法国;marine.goujon@univ-lille.fr (MG);justine.woszczyk@gmail.com (JW);kelliii@hotmail.fr (KG);thomas.sw@hotmail.fr (TS);sandy.fellah@univ-lille.fr (SF);jeanbaptiste.gibier@chru-lille.fr (J.-BG);isabelle.vanseuningen@inserm.fr (IVS);romain.larrue@univ-lille.fr (RL);christelle.cauffiez@univ-lille.fr (CC);viviane.gnemmi@chru-lille.fr (VG); sebastien.aubert@chru-lille.fr (SA); nicolas.pottier@univ-lille.fr (NP) 2 CHU Lille, Service d'Anatomo-Pathologie, F-59000 Lille, France 3 CHU Lille, Service de Toxicologie et Génopathies, F-59000 Lille, France * 通讯地址:michael.perrais@inserm.fr;电话:+33-3-20-29-88-62 † 这些作者对这项工作做出了同样的贡献。

电子电路 II – SECA1401
理想放大器将提供稳定的输出,该输出是输入信号的放大版本。但是,由于设备参数变化或环境温度变化以及设备的非线性,实际放大器的增益和稳定性并不是很好。可以通过反馈技术避免此问题,其中将输出信号的一部分反馈到输入并与输入信号相结合以产生所需的输出。反馈可以是负的(负反馈)或正的(正反馈)。在负反馈中,输出信号的一部分从输入信号中减去,在正反馈中,输出信号的一部分添加到输入信号以产生所需的输出。负反馈在几乎所有放大器偏置电路的稳定中都起着非常重要的作用,它使静态点的位置变得稳定。因此,它可以保持放大器增益值不受温度变化、电源电压等的影响。反馈可分为两种类型。



2022; 18(11): 4497-4512。doi: 10.7150/ijbs.73428 研究论文 烟雾诱导的 SAV1 基因启动子高甲基化破坏 YAP 负反馈和 P
YAP (基因符号YAP1) 作为一种潜在的致癌蛋白,与多种肿瘤的恶性程度呈正相关。然而,在多种正常组织细胞中单独过表达YAP并不能诱导肿瘤形成,其潜在机制尚不清楚。本文表明,YAP激活直接诱导其负调节因子SAV1的转录构成负反馈回路,该回路在维持肺上皮细胞稳态中起着至关重要的作用,在非小细胞肺癌(NSCLC)中失调。值得注意的是,吸烟促进SAV1启动子区域的高甲基化,通过失活Hippo通路破坏YAP负反馈。此外,外源性过表达SAV1可以充当运输蛋白,激活Hippo信号并同时抑制WNT通路以减少癌细胞生长。此外,利用肺癌类器官,我们发现慢病毒介导的 SAV1 基因转移联合甲基化抑制剂和 YAP-TEAD 抑制剂是肺癌患者(尤其是吸烟人群)的潜在可行临床用药方案。因此,这种 SAV1 介导的反馈回路提供了一种有效的机制来建立 YAP 调控的稳健性和稳态,并可作为吸烟 NSCLC 人群基因治疗的潜在靶点。

电子器件与电路(规
了解正反馈和负反馈系统所需的功能。 UNIT I PN 结器件 9 PN 结二极管 – 结构、操作和 VI 特性、扩散和过渡电容 - 削波和钳位电路 - 整流器 – 半波和全波整流器 – 显示设备 - LED、激光二极管、齐纳二极管特性 - 齐纳反向特性 – 齐纳作为稳压器 UNIT II 晶体管和晶闸管 9 BJT、JFET、MOSFET – 结构、操作、特性和偏置 UJT、晶闸管和 IGBT - 结构和特性。 UNIT III 放大器 9 BJT 小信号模型 – CE、CB、CC 放大器分析 – 增益和频率响应 – MOSFET 小信号模型 – CS 和源极跟随器分析 – 增益和频率响应单元 IV 多级放大器和差分放大器 9 BIMOS 级联放大器、差分放大器 – 共模和差模分析 – FET 输入级 – 单调谐放大器 – 增益和频率响应 – 中和方法、功率放大器 – 类型(定性分析)。单元 V 反馈放大器和振荡器 9 负反馈的优点 – 电压/电流、串联、并联反馈 – 正反馈 – 振荡条件、相移 – 维恩电桥、哈特利、考毕兹和晶体振荡器。
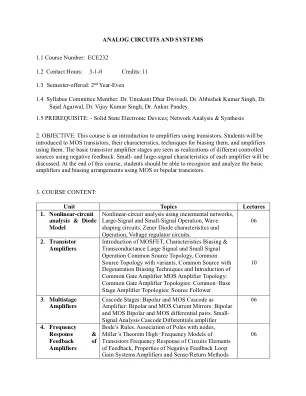
模拟电路和系统1.1课程编号
2。目的:本课程是使用晶体管的放大器的介绍。学生将被介绍给MOS晶体管,其特征,偏见的技术以及使用它们的放大器。基本的晶体管放大器阶段被视为使用负反馈对不同受控源的实现。每个放大器的小信号和大信号特征。在本课程结束时,学生应该能够使用MOS或双极晶体管识别和分析基本的放大器和偏置安排。