XiaoMi-AI文件搜索系统
World File Search System骨髓细胞

用户trecondi 1 g粉末的信息用于...
在开始使用此药物之前,请仔细阅读所有这些传单,因为它包含重要信息。•保留此传单。您可能需要再次阅读。•如果您还有其他问题,请询问您的医生,药剂师或护士。•如果您有任何副作用,请与您的医生,药剂师或护士交谈。这包括此传单中未列出的任何可能的副作用。请参阅第4节。此传单中的内容1。什么是trecondi,以及2。在给予trecondi 3.如何使用trecondi 4。可能的副作用5。如何存储trecondi 6。包装和其他信息的内容1。什么是trecondi及其用于trecondi的是包含活性物质treosulfan,该物质属于一组称为烷基化剂的药物。treosulfan用于为患者准备骨髓移植(造血干细胞移植)。treosulfan破坏了骨髓细胞,并实现了新的骨髓细胞的移植,从而导致健康血细胞的产生。trecondi被用作成人,青少年和一个超过一个月的癌症和非癌性疾病的青少年和儿童的血管细胞移植之前的治疗方法。2。警告和预防措施Trecondi是一种用来减少血细胞数量的细胞(细胞毒性)药物。在建议的剂量下,这是所需的效果。您将在治疗期间进行定期的血液检查,以检查您的血细胞计数不会太低。在给予trecondi trecondi之前,您不得对trecondi不得不给您如果您对treosulfan过敏,如果您患有积极的不受控制的感染,如果您患有严重的心脏,肺,肝脏或肾脏疾病,如果您患有遗传性DNA修复疾病,则可以忍受car,如果您患有严重的心脏,肺,肝脏或肾脏疾病,则可以将其恢复到遗传性的状态。您怀孕了,或者认为您可能怀孕。为了预防和治疗感染,您将获得药物,例如抗生素,抗真菌性或抗病毒药。trecondi可能会增加未来患其他癌症的风险。

抑制肿瘤固有的banf1通过CGAS刺激激活抗肿瘤免疫反应,并增强PD-1阻滞的功效
抽象背景banf1众所周知是基因组自DNA的环状GMP-AMP合酶(CGA)活性的自然对手。然而,班夫1在肿瘤免疫中的作用尚不清楚。在这里,我们研究了Banf1对抗肿瘤免疫和对免疫疗法的反应的可能影响。方法分析了癌症基因组公共数据,以评估Banf1表达,患者的生存和免疫细胞浸润的相关性。我们监测了肿瘤的生长,并探索了靶向肿瘤 - 内膜班夫1与MC38或B16F10肿瘤模型中的抗编程细胞死亡蛋白1(PD-1)结合使用的抗肿瘤功效。流式细胞仪,免疫荧光和T细胞耗尽实验用于验证BANF1在肿瘤免疫微环境重编程中的作用。RNA测序来询问Banf1如何调节抗肿瘤免疫的机制。结果我们表明,肿瘤组织中BANF1的表达上调与存活不良显着相关,并且与免疫细胞浸润呈负相关。肿瘤细胞中BANF1的缺乏显着拮抗免疫能力但没有免疫功能低下的小鼠的肿瘤生长,并增强了黑色素瘤和结肠癌鼠模型中对免疫疗法的反应。在免疫疗法临床队列中,较高的BANF1表达患者的预后较差。结论BANF1是CGAS刺激途径介导的抗肿瘤免疫力的关键调节剂。机械上,BANF1敲除激活由干扰素基因(CGAS-Sting)途径的CGAS-合酶刺激剂介导的抗肿瘤免疫反应,从而导致免疫激活的肿瘤微环境,包括增加的CD8 + T细胞浸润和减少的骨髓细胞诱导的细胞富含骨髓细胞的富含抑制剂。因此,我们的研究提供了一种理性的,即靶向Banf1是增强BANF1上调的癌症免疫疗法的有效策略。

引文 Brunetti M, Andersen K, Trøen G, Micci F, Spetalen S, Lenartova A, Tandsæther MR 和 Panagopoulos I (2024) 分子遗传表征
在 Xq13 带处发生断裂和重新连接的等着丝粒染色体 idic(X)(q13) 和 X 染色体长臂上的等染色体 i(X)(q10) 是癌症中罕见的细胞遗传学异常 ( 1 , 2 )。“ Mitelman 癌症染色体畸变和基因融合数据库 ”( 1 ) 的最新更新(2024 年 4 月 15 日)包含 47 个携带 idic(X)(q13) 的条目和 55 个携带 i(X)(q10 ) 的条目。idic (X)(q13) 主要见于被诊断为骨髓增生异常综合征 (MDS) 或急性髓细胞白血病 (AML) 的老年女性,在大多数情况下通常是唯一的细胞遗传学畸变 ( 1 , 3 – 8 )。相反,在各种肿瘤,包括 MDS 和 AML ( 1 ) 的复杂核型中,i(X)(q10) 多为继发性畸变。在 AML 和 MDS 的个案中,i(X)(q10) 是唯一的细胞遗传学异常 ( 9 , 10 )。仅在少数 MDS/AML 病例中报道了 Xq13 带中基因组断点的详细描述 ( 5 , 11 , 12 )。还发现患有 idic(X)(q13) 的 MDS/AML 患者的骨髓细胞中携带额外的亚微观遗传畸变 ( 5 , 13 )。尚未报道对 i(X)(q10 ) 病例中可能存在的其他遗传畸变进行调查。i(X)(q10) 的主要后果被认为是 Xp 的丢失和 Xq 上几个基因的获得。此外,其他遗传异常,包括 Tet 甲基胞嘧啶双加氧酶 2 ( TET2 ) 基因的致病变异,已被认为是 idic(X) 阳性髓系恶性肿瘤患者的常见继发事件 ( 5 )。由于携带 idic(X) (q13) 或 i(X)(q10) 的髓系肿瘤罕见,且对其致病机制的了解尚不完全,我们在此介绍了五种髓系肿瘤的分子细胞遗传学和致病变异的特征

全局DNA甲基化和多发性骨髓瘤患者的DNMT3A表达增加
目标。本研究的目的是比较多发性骨髓瘤(MM)不同阶段的患者中选定的DNA甲基转移酶和全球DNA甲基化状态的表达谱。进行分析,使用了不同的细胞群,包括未分离的骨髓瘤细胞和一组用相关抗体纯化的浆细胞。因此,将实验室数据与患者的临床数据进行了比较。患者和方法。进行分析,使用了44例44例患者(30例新诊断,9例复发和5例缓解患者)的未排序骨髓细胞群,并使用了8例患者的样本样本。我们使用了从3个健康个体的BM中分离出的COM可用RNA作为对照样品。通过定量RT-PCR进行了三种DNA甲基转移酶 - DNMT1,DNMT3A和DNMT3B的表达分析,并通过比色测定法检测到患者全局DNA甲基化谱。结果。未改变的DNMT1表达。归一化的DNMT3A基因表达在全球范围内更高。低(0.08–1.81%)的全球DNA甲基化状态在多发性骨髓瘤患者的无分类样品中与受监测的DNA甲基转移酶的SION谱或基于Durie-Salmon和International分期系统的MM阶段相关。结论。这是多发性骨髓瘤患者的不同阶段中DNA甲基转移酶表达谱和全局DNA甲基化状态之间的首次比较研究。注册了多发性骨髓瘤患者的未分布细胞群之间的全球甲基化水平与临床阶段之间没有显着相关性。DNMT3A基因的过表达发生在多发性骨髓瘤的患者的分类细胞和未分布的细胞群中。这一事实强调了DNMT3A是多发性骨髓瘤肿瘤前体的潜在标记。此外,我们在分类细胞群体和未分类的细胞群体中表达了DNA甲基转移酶表达的可比结果。这是从有条理的角度来看的一个有希望的结果,因为与未分类的多个骨髓瘤细胞样品相比,分类细胞的样品会减少所执行的可能分析的数量。

5月20日,星期一
P1。 Bernadette Tiberi HDAC7对于造血干和祖细胞功能Thomas Jefferson University P2是必需的。 greta zara lps介导的严重炎症重定向骨髓造血干细胞循环和分化命运,通过在希望城市贝克曼研究所P3上重塑其染色质结构。 Brandon T. Tran的骨髓细胞和祖细胞的表观遗传分析鉴定了细胞类型和基因靶标在HSPC训练有素的免疫中至关重要。 贝勒医学院P4。 wantong li解码转录因子依赖性增强子基因调节网络定义造血生态位功能。 俄亥俄州立大学P5。 RNA甲基化景观的单细胞和高分辨率映射 lla甲基化景观的高分辨率图显示了不列颠哥伦比亚省P6的造血干/祖细胞标识大学的表转录特征。 Monica kasbekar正常和美质前的人类HSC表现出对IL-1β哥伦比亚干细胞启动P7的年龄依赖性反应。 Xuan Zhang人类造血祖细胞的多模式地图:对辛辛那提儿童医院医疗中心P8的健康,衰老和疾病的见解。 詹姆斯·斯旺(James Swann)缺乏TET2的造血干和祖细胞中的表观遗传扰动会导致紧急骨髓骨髓疾病哥伦比亚大学P9。 Tanner C. Martinez Cux1通过调节芝加哥大学医学综合癌症中心P10来控制HSC命运。 Mona Vogel葡萄糖保留通过补体成分C3的细胞内水平调节HSC功能。P1。Bernadette Tiberi HDAC7对于造血干和祖细胞功能Thomas Jefferson University P2是必需的。greta zara lps介导的严重炎症重定向骨髓造血干细胞循环和分化命运,通过在希望城市贝克曼研究所P3上重塑其染色质结构。Brandon T. Tran的骨髓细胞和祖细胞的表观遗传分析鉴定了细胞类型和基因靶标在HSPC训练有素的免疫中至关重要。贝勒医学院P4。wantong li解码转录因子依赖性增强子基因调节网络定义造血生态位功能。俄亥俄州立大学P5。RNA甲基化景观的单细胞和高分辨率映射 lla甲基化景观的高分辨率图显示了不列颠哥伦比亚省P6的造血干/祖细胞标识大学的表转录特征。 Monica kasbekar正常和美质前的人类HSC表现出对IL-1β哥伦比亚干细胞启动P7的年龄依赖性反应。 Xuan Zhang人类造血祖细胞的多模式地图:对辛辛那提儿童医院医疗中心P8的健康,衰老和疾病的见解。 詹姆斯·斯旺(James Swann)缺乏TET2的造血干和祖细胞中的表观遗传扰动会导致紧急骨髓骨髓疾病哥伦比亚大学P9。 Tanner C. Martinez Cux1通过调节芝加哥大学医学综合癌症中心P10来控制HSC命运。 Mona Vogel葡萄糖保留通过补体成分C3的细胞内水平调节HSC功能。lla甲基化景观的高分辨率图显示了不列颠哥伦比亚省P6的造血干/祖细胞标识大学的表转录特征。Monica kasbekar正常和美质前的人类HSC表现出对IL-1β哥伦比亚干细胞启动P7的年龄依赖性反应。Xuan Zhang人类造血祖细胞的多模式地图:对辛辛那提儿童医院医疗中心P8的健康,衰老和疾病的见解。詹姆斯·斯旺(James Swann)缺乏TET2的造血干和祖细胞中的表观遗传扰动会导致紧急骨髓骨髓疾病哥伦比亚大学P9。Tanner C. Martinez Cux1通过调节芝加哥大学医学综合癌症中心P10来控制HSC命运。Mona Vogel葡萄糖保留通过补体成分C3的细胞内水平调节HSC功能。shorichiro takeishi造血干细胞数不完全由利基可用性阿尔伯特·爱因斯坦医学院和露丝·L·露丝·戈特斯曼(Ruth L.)和大卫·戈特斯曼(David S.分子医学研究所ULM大学和辛辛那提儿童医学中心
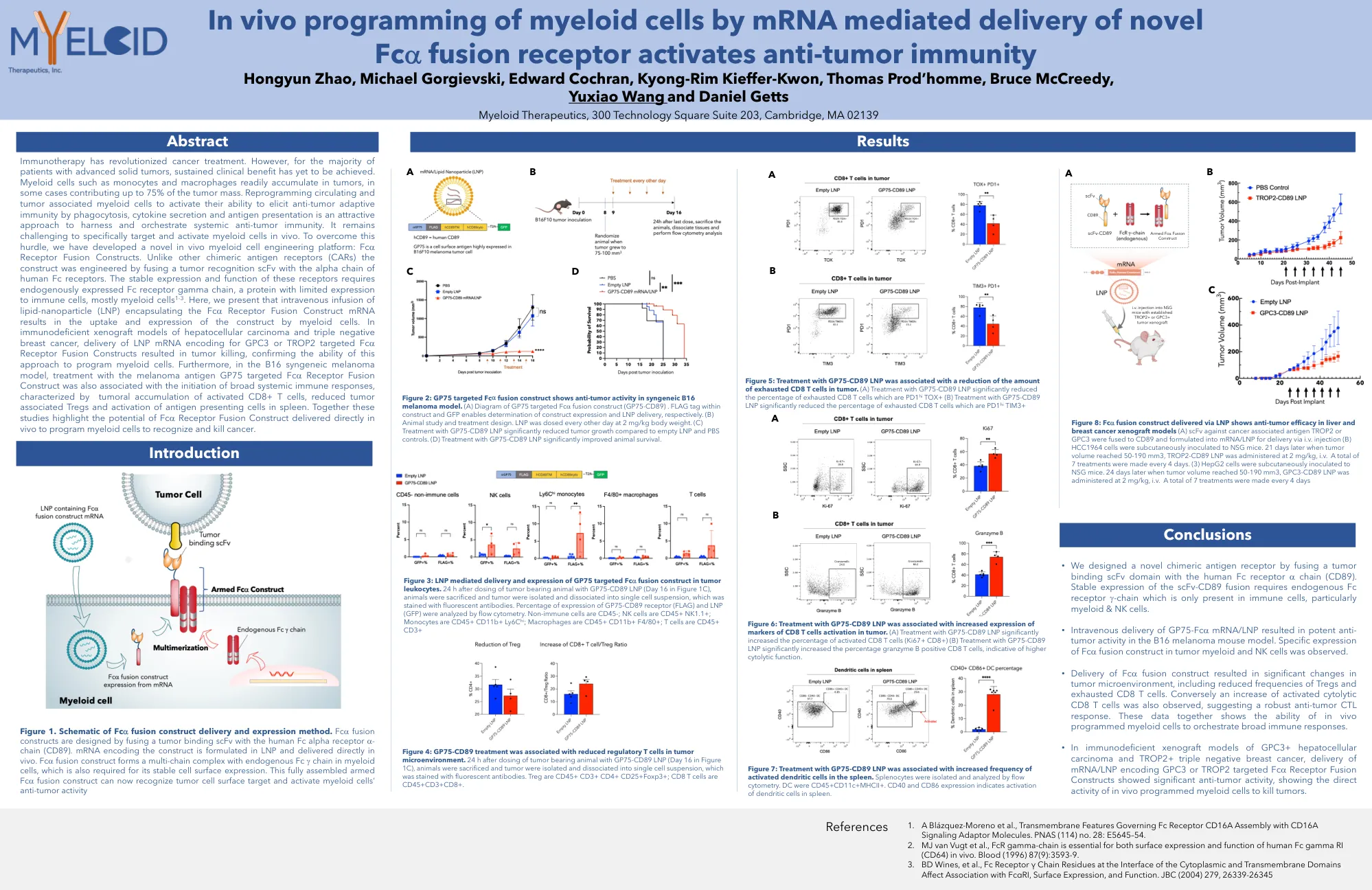
AACR2022体内递送海报
免疫疗法彻底改变了癌症治疗。但是,对于大多数晚期实体瘤患者,尚未实现持续的临床益处。髓样细胞(如单核细胞和巨噬细胞)很容易积聚在肿瘤中,在某些情况下,肿瘤质量的75%。重编程循环和肿瘤与髓样细胞相关,以激活其通过吞噬作用,细胞因子分泌和抗原表现来激活抗肿瘤适应性免疫的能力,是一种有吸引力的方法,可利用并策划系统性的抗肿瘤免疫。在体内专门靶向和激活髓样细胞仍然具有挑战性。为了克服这一障碍,我们开发了一种新型的体内髓细胞工程平台:FC A受体融合构建体。与其他嵌合抗原受体(CAR)不同,该构建体是通过将肿瘤识别SCFV与人体FC受体的α链融合而设计的。这些受体的稳定表达和功能需要内源表达的FC受体γ链,FC受体γ链是一种对免疫细胞表达有限的蛋白质,主要是髓样细胞1-3。在这里,我们介绍了包裹FC A受体融合构建体mRNA的静脉输注脂质 - 纳米颗粒(LNP)导致髓样细胞对构建体的摄取和表达。在肝细胞癌和三重阴性乳腺癌的免疫缺陷异种移植模型中,针对GPC3或trop2靶向FC的LNP mRNA的递送A受体融合构建体导致肿瘤杀死,从而确认了这种方法为骨髓细胞编程的能力。此外,在B16合成性黑色素瘤模型中,用黑色素瘤抗原GP75靶向FC A受体融合构建体的治疗也与启动广泛的全身免疫反应的启动有关,其特征在于肿瘤积累活化的CD8+ T细胞,可减少与肿瘤相关的TREG和SpleeNing spleen and spleen spleen and spleen的活化。这些研究共同强调了FC A受体融合构建体的潜力,直接在体内传递以编程髓样细胞以识别和杀死癌症。

脐带血库:平衡干细胞疗法中的炒作和希望
近年来,Stem-Cell疗法已成为一个有前途且高级的研究主题。治疗方法的发展产生了巨大的期望。使用干细胞的广泛可能性使这种尖端疗法成为现代医学的转折点,并为无法治愈的疾病提供希望1。脐带血是生物学材料,在出生和切割脐带后仍保留在脐带和胎盘中。它是具有高增殖潜力的干细胞的宝贵来源。尽管在许多国家,直到最近,它被视为医疗废物,并与胎盘和脐带一起处置,但现在经常收集它以隔离干细胞以存放并随后使用。脐带血是造血干细胞的丰富来源,可以像骨髓细胞一样被移植以重建造血和免疫系统。它们显示出比从成年供体获得的干细胞2。干细胞是人体中非专科细胞,其优势之一是它们无限期地自我更新的能力,从而保持体内恒定的细胞群体水平。这些细胞还可以分化为专业的祖细胞类型。由于这一过程,他们采用了执行专业功能所必需的形态和生化特性。从未分化状态到完全分化状态的过渡是逐渐的1。在1988年进行第一次移植后,脐带血已成为造血干细胞的标准来源3,4。与它们的分化能力有关,可以将干细胞分为以下组:1)整数细胞,该细胞源自胚泡,并具有分化为任何细胞类型的能力; 2)多能细胞,可以发展成除生殖细胞以外的任何细胞; 3)一项单位,包括仅分化为一种细胞类型的前体细胞)。关于起源,有三种类型的干细胞,即胚胎干细胞(全能或多能),在成人生物体的器官(多能或独立性)的器官中发现的体细胞,以及来自脐带血的干细胞2。在2006年,欧洲骨髓移植组从脐带血液中等同于骨髓骨髓和造血干细胞,从骨髓和rhu-g-csf动员后的外周血等同。自那时以来,无论其来源4,使用造血干细胞用于移植目的的适应症一直保持不变。本社论强调了脐带血库的重要性,并强调了知识在决策中的关键作用。对脐带血液库的信息良好的妇女可以更好地做出与自己的需求和家庭的需求相符的明智选择。通过强调教育和意识的重要性,这部社论旨在使妇女充满信心和清晰地驾驶脐带血液库的复杂性。

数据表:MCA1396D550产品详细信息
参考文献1。Els Conrath,K。等。(2001)骆驼单域抗体作为双特异性和双价抗体构建体中的模块化建筑单元。J Biol Chem。 276(10):7346-50。 2。 suen,J.L。 等。 (2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。 免疫。 103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。276(10):7346-50。2。suen,J.L。等。(2001)NZB x NZW F 1小鼠中骨髓衍生的树突状细胞对U1a蛋白的自动T细胞反应和U1A蛋白的抗原决定因素的表征。免疫。103:301-309。 3。 Hoffmann,S.C。等。 (2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。103:301-309。3。Hoffmann,S.C。等。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。 J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。(2007)鉴定Clec12b,骨髓细胞上的抑制性受体。J Biol Chem。 282(31):22370-5。 4。 Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。282(31):22370-5。4。Zheng,J。等。 (2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。 BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Zheng,J。等。(2007)在Treg抑制作用中免疫的小鼠的血清将DEK鉴定为神经母细胞瘤肿瘤抗原。BMC免疫。 8:4。 5。 Bahi,A。 &Dreyer,J.L。 (2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。 基因脑行为。 7(2):244-56。 6。 Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。BMC免疫。8:4。5。Bahi,A。&Dreyer,J.L。(2008)伏伏核中纤溶酶原激活剂的过表达增强了可卡因,苯丙胺和吗啡诱导的奖励和行为敏化。基因脑行为。7(2):244-56。6。Wrighton,K.H。 等。 (2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。 J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。Wrighton,K.H。等。(2009)转化生长因子{beta}可以独立于骨形态发生蛋白受体刺激Smad1磷酸化。J Biol Chem。 284(15):9755-63。 7。 Diefenbacher,M。等。 (2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。 8。J Biol Chem。284(15):9755-63。7。Diefenbacher,M。等。(2011)DSL1绑扎复合物积极参与可溶性NSF(N-乙基甲米酰胺敏感因子)附着蛋白受体(SNARE)复合物组件在塞雷维氏酵母中的内质网处。8。J Biol Chem。 286:25027-38。 Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放J Biol Chem。286:25027-38。Alvarez,M.M。 等。 (2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。 PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Alvarez,M.M。等。(2010)人血清中流感A/H1N1/2009抗体的特定识别:一种简单的无病毒ELISA方法。PLOS ONE。 5:E10176。 9。 Bahi,A。等。 (2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。 神经心理药理学。 33:2726-34。 10。 Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放PLOS ONE。5:E10176。9。Bahi,A。等。(2008)组织型纤溶酶原激活剂系统在苯丙胺诱导的条件位置偏好灭绝和恢复中的作用。神经心理药理学。33:2726-34。10。Gunnarsen,K.S。 等。 (2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。 BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Gunnarsen,K.S。等。(2010)可溶性单链T细胞受体的周质表达由伴侣FKPA挽救。BMC生物技术。 10:8。 11。 Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放BMC生物技术。10:8。11。Hwang,H.Y。 等。 (2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。 mol免疫。 45:2570-80。 12。 de Vooght,L。等。 (2012)功能性的表达和细胞外释放Hwang,H.Y。等。(2008)高度特异性的抑制C1Q球形与人IgG结合:一种使用工程的单链抗体可变片段来控制和调节经典补体途径的新方法。mol免疫。45:2570-80。12。de Vooght,L。等。(2012)功能性的表达和细胞外释放

2021 年 ASPHO 会议论文和海报索引
全体会议论文 # 2001 黏连蛋白功能改变对核心结合因子急性髓系白血病增殖的影响 Shannon Conneely、Jason Rogers、Matthew Miller、Jason Guo、Rohit Gupta、Geraldo Medrano、Debananda Pati、Rachel Rau 贝勒医学院/德克萨斯儿童医院,美国德克萨斯州休斯顿 背景:核心结合因子急性髓系白血病 (AML) 是一种常见的儿童 AML,其特征是 inv(16) 或 t(8;21) 病变,这些病变会抑制核心结合因子复合物的功能。尽管这些重排被认为是 AML 的有利风险,但近 30% 的核心结合因子 AML 儿童会复发,这表明需要继续加深对 AML 生物学的了解和寻找新的治疗靶点。黏连蛋白复合体基因突变常见于 t(8;21) AML,但在 inv(16) AML 中从未发现,这表明黏连蛋白在每种核心结合因子 AML 亚型的病理生理学中发挥着独特的作用。目标:本项目的目标是确定黏连蛋白突变如何改变核心结合因子 AML 的生物学特性。我们假设,黏连蛋白正常功能的丧失会增强表达 t(8;21) AML 特征性 RUNX1-CBFA2T1 (RC) 融合蛋白的细胞增殖,并抑制表达 inv(16) AML 特征性 CBFß-SMMHC (CS) 融合的细胞的增殖能力。设计/方法:从黏连蛋白正常 (Smc3 +/+) 或黏连蛋白单倍体不足 (Smc3 +/-) 的小鼠体内采集骨髓细胞。我们利用逆转录病毒转导来表达空载体对照、RC 融合或 CS 融合蛋白。然后将转导的细胞接种在含有干细胞和骨髓促进细胞因子的甲基纤维素中,进行连续接种试验,或移植到致死性辐射受体小鼠体内,以评估对白血病转化的影响。结果:连续接种试验表明,黏连蛋白单倍体不足会增加表达 RC 蛋白的细胞的集落形成能力,并降低表达 CS 蛋白的细胞的集落形成能力。黏连蛋白单倍体不足会改变几种关键造血调节基因的表达,尽管这些影响取决于存在哪种融合蛋白。在小鼠 RC 模型中,无论黏连蛋白功能如何,都会发展为未分化白血病。然而,二次移植模型显示,黏连蛋白功能下降会导致白血病存活时间缩短,骨髓浸润增加。结论:正常黏连蛋白功能的丧失对表达核心结合因子 AML 融合蛋白的细胞增殖有不同的影响。在表达与 t(8;21) AML 相关的 RC 融合的细胞中,黏连蛋白功能的降低在白血病转化之前提供了生长优势,并带来了更具浸润性和侵袭性的白血病表型。或者,黏连蛋白功能下降导致表达 inv(16) AML CS 融合的细胞生长不利,造血基因表达发生显著变化。未来的实验将重点阐明核心结合因子 AML 中黏连蛋白功能下降所改变的潜在细胞机制。