XiaoMi-AI文件搜索系统
World File Search System高含量

用于纺织对称电容器的 PEDOTS:PSS@KNF 线状电极
代表着一种更可靠、更安全、生命周期更长的替代方案。通过湿纺技术成功获得了许多由石墨烯、碳纳米管、导电聚合物以及最近的 MXenes 制成的纤维,并研究将其作为可穿戴超级电容器的一维电极。[17–29] 然而,这些材料通常涉及复杂的合成程序、有害的分散剂溶剂或后处理步骤,以生产出具有足够机械阻力和电化学性能的纤维。芳族聚酰胺纳米纤维 (ANF) 最近被提议作为一种新的纳米级构建块来设计新的复合材料。[30] 与基于单体聚合的标准路线相反,ANF 可以通过自上而下的方法轻松快速地获得,通过溶解芳族聚酰胺聚合物链,然后通过溶液加工重新组装成宏观纤维或薄膜。[30,31] 芳族聚酰胺聚合物以其机械强度而闻名,但它不导电,必须负载导电填料才能实现电子传输。到目前为止,ANF 主要被研究用作聚合物增强体的填料[32,33]、多功能膜的基质[34–37]、隔热罩[38,39],甚至用作隔膜的添加剂和锂离子电池的固态电解质。[40,41] 然而,尽管 KNF 分散体具有良好的湿纺性,但人们对使用 ANF 来制造 FSC 却关注甚少。在之前的工作中,Cao 等人通过共湿纺核碳纳米管分散体和鞘 ANF 分散体制备了具有核壳结构的纤维。[42] 通过用 H3PO4/PVA 凝胶电解质渗透获得的对称 FSC 显示出高达 0.75 mF cm −1 的显著线性容量。Wang 等人将石墨烯纳米片 (GNPs) 加载到 ANF 分散体中,通过在水/乙酸溶液中凝固获得 ANFs/GNPs 复合线状电极。[43] 然而,他们的结果表明,GNPs 通过恢复对苯二甲酰胺单元之间的氢键干扰了 ANFs 的凝固,导致在 ANFs 基质中 GNPs 高含量时拉伸强度持续下降。在这项工作中,PEDOT:PSS@KNFs 复合纤维通过一个简单的两步工艺生产出来,包括将 Kevlar 纳米纤维化为 Kevlar 纳米纤维 (KNF)、KNF 纤维的湿纺以及随后浸泡在 PEDOT:PSS 水分散体中。以这种方式,由于导电的 PEDOT:PSS 链渗透而几乎保持 KNF 基质的机械阻力不变,因此获得了导电纤维。 PEDOT:PSS@KNF 纤维具有柔韧性、可编织、可缝纫等特点,通过耦合相邻的两根纤维,可以形成对称的 FSC。

在撒哈拉以南非洲的肥料使用:类型和金额
在撒哈拉以南非洲的肥料使用:类型和金额v.a.Kelly和A. Naseem农业经济学系,密歇根州立大学,美国关键词:肥料,撒哈拉以南非洲,土壤退化,内容1。 萨哈拉以南非洲的肥料使用历史1.1。 背景1.2肥料消耗1.3。 肥料强度使用1.4。 使用的肥料类型2. 影响肥料增长的因素2.1历史和政策影响2.2。 农业生态区和其他地理因素2.3。 殖民遗产2.4。 人口统计2.5。 国民收入2.6。 基础设施2.7。 作物选择2.8价格和盈利能力3。 化肥对作物生产和环境的影响4。 未来的趋势预期的致谢词汇表书记传记草图总结了撒哈拉以南非洲肥料使用的显着特征(SSA)是(1)使用的有机和无机肥料的数量极少,使用的有机和无机肥料,(2)非常低的强度(2)使用的使用速度很低(Kg/ha),以及(3)的肥料速率。 虽然世界上许多地区都正确关注过度肥料使用的负面影响(富营养化,盐水,铝毒性等。 SSA中的总肥料消耗可能会在未来五到十年的时间内继续缓慢增长,而在消费量的年间波动中仍然很重要Kelly和A. Naseem农业经济学系,密歇根州立大学,美国关键词:肥料,撒哈拉以南非洲,土壤退化,内容1。萨哈拉以南非洲的肥料使用历史1.1。背景1.2肥料消耗1.3。肥料强度使用1.4。使用的肥料类型2.影响肥料增长的因素2.1历史和政策影响2.2。农业生态区和其他地理因素2.3。殖民遗产2.4。人口统计2.5。国民收入2.6。基础设施2.7。作物选择2.8价格和盈利能力3。化肥对作物生产和环境的影响4。未来的趋势预期的致谢词汇表书记传记草图总结了撒哈拉以南非洲肥料使用的显着特征(SSA)是(1)使用的有机和无机肥料的数量极少,使用的有机和无机肥料,(2)非常低的强度(2)使用的使用速度很低(Kg/ha),以及(3)的肥料速率。虽然世界上许多地区都正确关注过度肥料使用的负面影响(富营养化,盐水,铝毒性等。SSA中的总肥料消耗可能会在未来五到十年的时间内继续缓慢增长,而在消费量的年间波动中仍然很重要),大多数SSA的使用太少了肥料的负面影响(土壤养分的迅速损失,生物量不足,用于回收养分和土壤有机物,由于培养转移的培养,将培养扩大到高含量的土地等,森林和林地的丧失,造成了高度的土地等)。
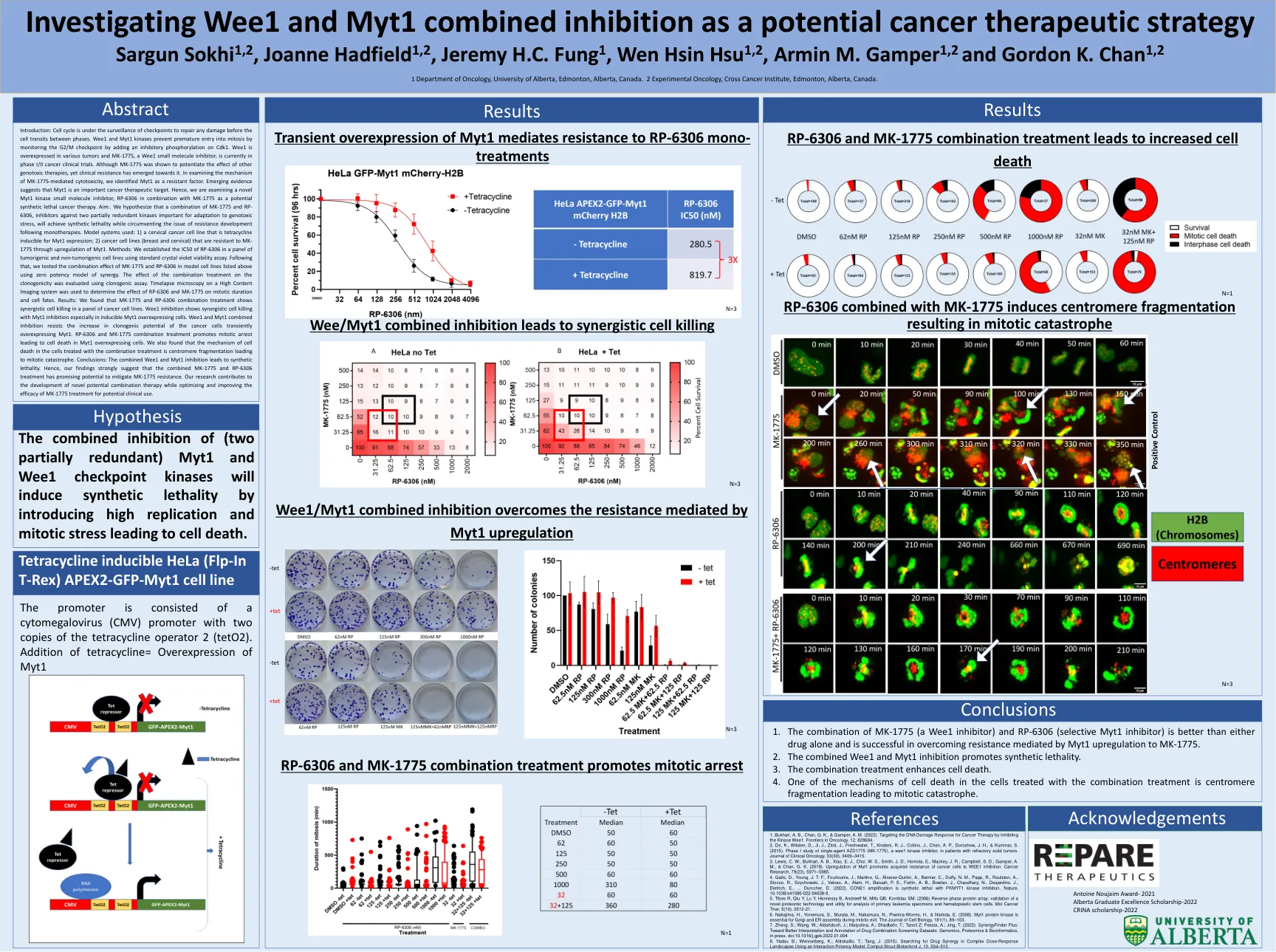
研究WEE1和MYT1作为潜在的癌症治疗策略Sargun Sokhi 1,2,Joanne Hadfield 1,2,Jeremy H.C.真菌1
简介:细胞周期处于检查点的监视下,以修复各个相之间的细胞过渡之前的任何损坏。WEE1和MYT1激酶通过在CDK1上添加抑制性磷酸化来监测G2/M检查点,以防止过早进入有丝分裂。WEE1在各种肿瘤中过表达,而MK-1775(WEE1小分子抑制剂)目前正在I/II期癌症临床试验中。尽管显示了MK-1775可以增强其他遗传毒性疗法的作用,但临床抗性却朝向其抗性。在检查MK-1775介导的细胞毒性的机理时,我们将MYT1确定为抗性因子。新兴证据表明,MYT1是重要的癌症治疗靶点。因此,我们正在研究一种新型的MYT1激酶小分子抑制剂RP-6306与MK-1775结合使用,作为潜在的合成致死性癌症治疗。aim:。我们假设MK-1775和RP-6306的组合是针对两个部分冗余激酶对适应遗传毒性应激很重要的抑制剂,将实现合成的致死性,同时绕过单一层次的耐药性发育问题。使用的模型系统:1)宫颈癌细胞系,是四环素可诱导的MyT1表达; 2)通过上调myt1,对MK-1775具有抗性的癌细胞系(乳房和颈椎)。方法:我们使用标准的晶体紫罗兰色生存力测定法在一组肿瘤和非肿瘤细胞系中建立了RP-6306的IC50。之后,我们使用协同效力模型在上面列出的模型细胞系中测试了MK-1775和RP-6306的组合效应。使用克隆生成测定法评估了组合处理对克隆原性的影响。对高含量成像系统上的时间段显微镜用于确定RP-6306和MK-1775对有丝分裂持续时间和细胞命运的影响。结果:我们发现MK-1775和RP-6306组合处理表明,癌细胞系中的协同细胞杀死。WEE1抑制作用显示了通过MYT1抑制的协同细胞杀死,尤其是在诱导的MYT1过表达细胞中。 WEE1和MYT1结合抑制作用可抵抗癌细胞瞬时过表达MYT1的克隆发育潜力的增加。 RP-6306和MK-1775组合治疗促进有丝分裂停滞,导致MYT1过表达细胞的细胞死亡。 我们还发现,用联合处理处理的细胞中细胞死亡的机理是丝粒碎片化导致有丝分裂灾难。 结论:合并的WEE1和MYT1抑制导致合成致死性。 因此,我们的发现强烈表明,MK-1775和RP-6306的组合治疗具有减轻MK-1775耐药性的潜力。 我们的研究有助于开发新型的潜在组合疗法,同时优化和提高MK-1775治疗的潜在临床用途的功效。WEE1抑制作用显示了通过MYT1抑制的协同细胞杀死,尤其是在诱导的MYT1过表达细胞中。WEE1和MYT1结合抑制作用可抵抗癌细胞瞬时过表达MYT1的克隆发育潜力的增加。RP-6306和MK-1775组合治疗促进有丝分裂停滞,导致MYT1过表达细胞的细胞死亡。我们还发现,用联合处理处理的细胞中细胞死亡的机理是丝粒碎片化导致有丝分裂灾难。结论:合并的WEE1和MYT1抑制导致合成致死性。因此,我们的发现强烈表明,MK-1775和RP-6306的组合治疗具有减轻MK-1775耐药性的潜力。我们的研究有助于开发新型的潜在组合疗法,同时优化和提高MK-1775治疗的潜在临床用途的功效。

与针对2型炎症的生物学疗法相比,JAK抑制剂治疗期间TARC的差异动力学
1。Catherine J,Roufosse F.升高的TARC/CCL17挤压告诉我们有关嗜酸性疾病的信息?semin免疫病。2021; 43(3):439-458。2。Renert-Yuval Y,Thyssen JP,Bisonette R等。特应性皮炎的生物标志物 - 代表国际湿疹委员会的综述。J过敏临床免疫。 2021; 147(4):1174-1190。 3。 Bakker DS,Ariens LFM,Giovanonne B等。 easi p-easi:使用血清生物标志物组合用dupilumab治疗的特应性皮炎患者进行预测的疾病严重程度。 过敏。 2020; 75(12):3287-3289。 4。 Olydam Ji,De Wijs Lem,Ropke MA等。 easi p-easi:预测用曲洛牛umab治疗的特应性皮炎患者的疾病严重程度。 J投资皮肤病。 2022; 142(12):3335-3337。 5。 Dubin C,Del Duca E,Guttman-Yassky E.特应性皮炎的IL-4,IL-13和IL-31途径。 专家Rev Clin Immunol。 2021; 17(8):835-852。 6。 Hagino T,Saeki H,Kanda N. upadacitinib治疗日本现实世界实践中中度至重度特应性皮炎的功效和安全性。 J Dermatol。 2022; 49(11):1158-1167。 7。 Gooderham M,Girolomoni G,Moore Jo等。 在2B期试验中停用治疗后中度至重度性皮炎患者对酰胺替尼的持久性。 皮肤病。 2022; 12(9):2077-2085。 8。 J过敏临床免疫。J过敏临床免疫。2021; 147(4):1174-1190。3。Bakker DS,Ariens LFM,Giovanonne B等。easi p-easi:使用血清生物标志物组合用dupilumab治疗的特应性皮炎患者进行预测的疾病严重程度。过敏。2020; 75(12):3287-3289。4。Olydam Ji,De Wijs Lem,Ropke MA等。 easi p-easi:预测用曲洛牛umab治疗的特应性皮炎患者的疾病严重程度。 J投资皮肤病。 2022; 142(12):3335-3337。 5。 Dubin C,Del Duca E,Guttman-Yassky E.特应性皮炎的IL-4,IL-13和IL-31途径。 专家Rev Clin Immunol。 2021; 17(8):835-852。 6。 Hagino T,Saeki H,Kanda N. upadacitinib治疗日本现实世界实践中中度至重度特应性皮炎的功效和安全性。 J Dermatol。 2022; 49(11):1158-1167。 7。 Gooderham M,Girolomoni G,Moore Jo等。 在2B期试验中停用治疗后中度至重度性皮炎患者对酰胺替尼的持久性。 皮肤病。 2022; 12(9):2077-2085。 8。 J过敏临床免疫。Olydam Ji,De Wijs Lem,Ropke MA等。easi p-easi:预测用曲洛牛umab治疗的特应性皮炎患者的疾病严重程度。J投资皮肤病。2022; 142(12):3335-3337。5。Dubin C,Del Duca E,Guttman-Yassky E.特应性皮炎的IL-4,IL-13和IL-31途径。专家Rev Clin Immunol。2021; 17(8):835-852。6。Hagino T,Saeki H,Kanda N. upadacitinib治疗日本现实世界实践中中度至重度特应性皮炎的功效和安全性。 J Dermatol。 2022; 49(11):1158-1167。 7。 Gooderham M,Girolomoni G,Moore Jo等。 在2B期试验中停用治疗后中度至重度性皮炎患者对酰胺替尼的持久性。 皮肤病。 2022; 12(9):2077-2085。 8。 J过敏临床免疫。Hagino T,Saeki H,Kanda N. upadacitinib治疗日本现实世界实践中中度至重度特应性皮炎的功效和安全性。J Dermatol。2022; 49(11):1158-1167。7。Gooderham M,Girolomoni G,Moore Jo等。在2B期试验中停用治疗后中度至重度性皮炎患者对酰胺替尼的持久性。皮肤病。2022; 12(9):2077-2085。8。J过敏临床免疫。J过敏临床免疫。Fujisawa T,Fujisawa R,Kato Y等。在血小板中存在高含量和激活调节的趋化因子,以及胸腺的元素水平以及激活调节的趋化因子和巨噬细胞衍生的趋化趋化因子的特征性皮肤钛患者。2002; 110(1):139-146。 9。 Kabashima K,Matsumara T,Komazaki H等。 nemolizumab加上特应性皮炎(AD)和中等症状瘙痒的患者的局部用药可改善瘙痒和AD迹象长达68周:两次III期长期研究的结果。 br j dermatol。 2022; 186(4):642-651。2002; 110(1):139-146。9。Kabashima K,Matsumara T,Komazaki H等。 nemolizumab加上特应性皮炎(AD)和中等症状瘙痒的患者的局部用药可改善瘙痒和AD迹象长达68周:两次III期长期研究的结果。 br j dermatol。 2022; 186(4):642-651。Kabashima K,Matsumara T,Komazaki H等。nemolizumab加上特应性皮炎(AD)和中等症状瘙痒的患者的局部用药可改善瘙痒和AD迹象长达68周:两次III期长期研究的结果。br j dermatol。2022; 186(4):642-651。

食物不吃糖尿病前
为防止糖尿病的发展,避免加剧前糖尿病的食物至关重要。诸如精制碳水化合物,甜谷物和加工肉类等食物增加了2型糖尿病和心脏病的风险。当血糖水平高于正常水平高但不足以分类为2型糖尿病时,就会发生糖尿病前期。通过改变生活方式的改变,包括饮食改造,例如避免某些食物,可以将血糖水平降低至健康范围。具有高血糖指数的精制碳水化合物被迅速消化,并导致血糖水平迅速增加。这些碳水化合物,包括白面包,米饭和面粉,缺乏纤维和必需的营养,导致食用后不久饥饿。饮食中高的碳水化合物的饮食增加了2型糖尿病,高血压和心脏病的风险。相反,应选择像全谷物,淀粉蔬菜,豆类和豆类等复杂的碳水化合物,因为它们的纤维较高并更慢地消化,提供持久的能量和饱腹感。甜糕点和甜点,高含糖,饱和脂肪和卡路里的高含量,由于它们与肥胖,2型糖尿病,炎症和心脏病的联系,应适度消耗。更健康的甜点选择包括新鲜的水果,搭配格兰诺拉麦片,全谷物吐司搭配坚果黄油和深色巧克力覆盖的杏仁。含糖的饮料是添加糖的主要来源,应由低糖选择取代,例如白糖,未加糖的茶或无糖调味水,以降低2型糖尿病,心脏病和中风的风险。1。而不是含糖饮料,选择茶,不添加糖的闪闪发光或低糖果汁。在谷物方面,请注意,热和冷甜味的选择通常很低,含糖量较高。取而代之的是,选择少于5克糖和至少3克纤维的全麦谷物,并在上面放上浆果,坚果或Chia种子,以增加营养。健康的早餐替代品包括带有火鸡香肠的炒鸡蛋,带有浆果的希腊酸奶,带鸡蛋的鳄梨吐司以及带坚果和新鲜水果的干酪碗。限制您对饱和脂肪的摄入,这可以降低胰岛素敏感性并有助于糖尿病的发展,并增加胆固醇水平和心脏病风险。取而代之的是,在鳄梨,脂肪鱼,橄榄油,花生酱,坚果和种子中发现了适量的健康单不饱和和多不饱和脂肪。加工的肉类,如培根,香肠和午餐肉的饱和脂肪和钠往往很高,并且与2型糖尿病,心脏病和癌症的风险增加有关。考虑将其代替植物性蛋白质来源,例如坚果,种子,小扁豆,无皮肤家禽和海鲜。在水果方面,最好的选择是最好的,因为干燥可以浓缩营养和糖含量。食用干果时要注意份量,并考虑避免使用糖尿病前期的血糖指数高的水果。未加糖的希腊酸奶是一种健康的零食选择,但要警惕含糖的浇头和调味料。上面放有坚果,种子和浆果,以增加风味。糖尿病前饮食需要平衡碳水化合物和蛋白质,以防止血糖峰值。希腊酸奶是一个不错的选择,糖和碳水化合物比传统酸奶少。还可以提供由杏仁,大豆或椰奶制成的低糖非奶油酸奶。由于碳水化合物含量高和血糖指数限制了炸薯条,这会导致血糖迅速升高。炸食品每周消耗3次,将2型糖尿病的风险增加近19%。选择烤的地瓜,欧洲防风草或小扁豆代替炸薯条。尝试脆皮羽衣甘蓝片,烤西葫芦,胡萝卜或绿豆“薯条”作为替代品。这些食物不会引起血糖峰值。烘烤炸薯条,与健康的脂肪和蛋白质配对也可以帮助保持血糖稳定。通过调味品改善食品风味对于糖尿病前饮食至关重要。但是,一些沙拉调味料和调味品,例如番茄酱,烧烤酱,蜂蜜芥末和法式调味料,含有高钠,碳水化合物,脂肪和卡路里。由于添加糖,许多无脂肪的敷料比常规版本多。选择用橄榄油,鳄梨油或其他醋制成的油性沙拉敷料是一种更健康的选择。而不是使用高糖果酱和果冻,而是制作低糖果酱或使用无糖果冻。用全谷物,健康脂肪,瘦蛋白,水果,蔬菜和豆类代替精制的碳水化合物,糕点,甜谷物和含糖饮料可以帮助管理血糖水平。匹兹堡,Papennsylvania 2。格林斯伯勒,NCNORTH CAROLINA 3。檀香山,hihawaii 8。美国的一些城市被列为有糖尿病前期的人,包括南卡罗来纳州的查尔斯顿;新泽西州帕特森;加利福尼亚州兰开斯特;田纳西州默弗里斯伯勒;加利福尼亚州圣罗莎;俄勒冈州尤金;路易斯安那州什里夫波特;田纳西州克拉克斯维尔;卡里,北卡罗来纳州;塞勒姆,俄勒冈州;弗吉尼亚州纽波特新闻;田纳西州查塔努加;俄亥俄州阿克伦;普罗维登斯,罗德岛;佛罗里达州塔拉哈西;亚利桑那州皮奥里亚;南达科他州苏福尔斯;纽约扬克斯;北卡罗来纳州费耶特维尔;纽约罗切斯特;得克萨斯州麦金尼;加利福尼亚州莫雷诺谷;加利福尼亚州弗里蒙特;德克萨斯州弗里斯科;华盛顿斯波坎;爱达荷州博伊西;佛罗里达州圣露西港;得克萨斯州欧文;温斯顿·塞勒姆(Winston-Salem),北卡罗来纳州;佛罗里达州圣彼得堡;德克萨斯州拉雷多;新泽西州泽西市;纽约布法罗;和密苏里州圣路易斯;亚利桑那州钱德勒;北卡罗来纳州达勒姆;内布拉斯加州林肯。圣保罗,Mnminnesota 4。 Newark,NJNew Jersey 5。 列克星敦,肯塔基6。 Stockton,Cacalifornia 7。 克利夫兰,俄亥俄州9。 阿灵顿,TXEXAS 10。 奥克兰,卡卡利尼亚11。 长滩,卡卡利尼亚12。 Raleigh,Ncnorth Carolina 13。 奥马哈,内内布拉斯加州14。 科罗拉多斯普林斯,cocolorado 15。 弗雷斯诺,cacalifornia 16。 Albuquerque,NMNEW墨西哥17。 孟菲斯,tntennessee 18。 华盛顿特区,华盛顿特区19。 拉斯维加斯,nvnevada 20。 西雅图,Wawashington圣保罗,Mnminnesota 4。Newark,NJNew Jersey 5。列克星敦,肯塔基6。Stockton,Cacalifornia 7。克利夫兰,俄亥俄州9。阿灵顿,TXEXAS 10。奥克兰,卡卡利尼亚11。长滩,卡卡利尼亚12。Raleigh,Ncnorth Carolina 13。 奥马哈,内内布拉斯加州14。 科罗拉多斯普林斯,cocolorado 15。 弗雷斯诺,cacalifornia 16。 Albuquerque,NMNEW墨西哥17。 孟菲斯,tntennessee 18。 华盛顿特区,华盛顿特区19。 拉斯维加斯,nvnevada 20。 西雅图,WawashingtonRaleigh,Ncnorth Carolina 13。奥马哈,内内布拉斯加州14。科罗拉多斯普林斯,cocolorado 15。弗雷斯诺,cacalifornia 16。Albuquerque,NMNEW墨西哥17。孟菲斯,tntennessee 18。华盛顿特区,华盛顿特区19。拉斯维加斯,nvnevada 20。西雅图,Wawashington西雅图,Wawashington
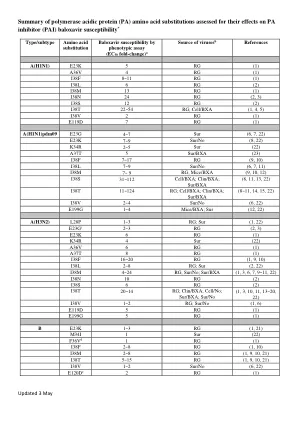
摘要 - 聚合酶 - 酸性 - (PA) - 蛋白质 - 氨基 -
* PA中的其他氨基酸取代,在参考文献1(Omoto S等,2018)和#2(Hashimoto T等,2020年)中研究了Baloxavir易感性没有变化的其他氨基酸取代。通过基于细胞培养的测定法评估(焦点,斑块或屈服分析,高含量成像中和(提示)和ViroDot分析)。EC 50倍变化。b细胞,细胞培养;临床试验;小鼠,鼠标模型; RG,反向遗传学; SUR,监视研究; BXA,在Baloxavir压力下选出的取代;不,Baloxavir不使用。c e23g(T0831)。通过表型测定测试了带有E23G的RG病毒。d对应于A36V A型A型PA中的A36V。 E对应于A型A型PA中的E119D。参考文献1。Omoto S,Speranzini V,Hashimoto T,Noshi T,Yamaguchi H,Kawai M,Kawaguchi K,Uehara T,Shishido T,Naito A,Naito A,Cusack S.2018。通过核酸内切酶抑制剂Baloxavir maroxil诱导的流感病毒变体的表征。SCI REP 8:9633。2。Hashimoto T,Baba K,Inoue K,Okane M,Hata S,Shishido T,Naito A,Wildum S,Omoto S.2020。在Baloxavir Marboxil的临床试验中检测到的流感病毒的三聚体RNA聚合酶复合物中氨基酸取代的全面评估。流感其他呼吸病毒DOI:10.1111/irv.12821。3。ince WL,Smith FB,O'Rear JJ,Thomson M.2020。J Infect DIS 222:957-961。 4。 2018。J Infect DIS 222:957-961。4。2018。治疗 - 伴随流感病毒聚合酶酸性取代率与Balosavir Maroxavir Marboxil试验中的i38中的i38中的酸性取代相关。Noshi T, Kitano M, Taniguchi K, Yamamoto A, Omoto S, Baba K, Hashimoto T, Ishida K, Kushima Y, Hattori K, Kawai M, Yoshida R, Kobayashi M, Yoshinaga T, Sato A, Okamatsu M, Sakoda Y, Kida H, Shishido T, Naito A.Baloxavir酸的体外表征,Baloxavir酸是一种流感病毒聚合酶PA亚基的第一类帽依赖性内切酶抑制剂。抗病毒Res 160:109-117。5。Takashita E,Morita H,Ogawa R,Nakamura K,Fujisaki S,Shirakura M,Kuwahara T,Kishida N,Watanabe S,Odagiri T.2018。流感病毒对新型帽依赖性核酸内切酶抑制剂baloxavir maroxil的敏感性。前微生物9:3026。6。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。 2019。 评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。 欧元监视24:1800666。 7。 Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A. 2020。 全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。 抗病毒Res 175:104718。 8。 2020。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。2019。评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。欧元监视24:1800666。7。Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A.2020。全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。抗病毒Res 175:104718。8。2020。Takashita E, Abe T, Morita H, Nagata S, Fujisaki S, Miura H, Shirakura M, Kishida N, Nakamura K, Kuwahara T, Mitamura K, Ichikawa M, Yamazaki M, Watanabe S, Hasegawa H, Influenza Virus Surveillance Group of J.流感A(H1N1)PDM09病毒,由于未经Baloxavir治疗的儿童检测到PA E23K替代而表现出对Baloxavir的敏感性降低。抗病毒Res 180:104828。 9。 Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。 2019。 亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。 抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 180:104828。9。Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。2019。亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 164:91-96。10。Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。2020。流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。Proc Natl Acad Sci U S A 117:8593-8601。11。Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。2020。季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。J Infect DIS 221:367-371。12。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y.2020。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。J Infect Dis 221:1699-1702。13。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。J Infect DIS 222:121-125。14。J Infect DIS 221:63-70。15。2020。16。在2018-2019流感季节治疗流感A的儿童后,检测Baloxavir Marboxil易感性降低的变体。Checkmahomed L,M'Hamdi Z,Carbonneau J,Venable MC,Baz M,Abed Y,Boivin G.2020。抗性抗性聚合酶酸I38T取代对当代流感A(H1N1)PDM09和A(H3N2)菌株的适应性的影响。Imai M, Yamashita M, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kiso M, Murakami J, Yasuhara A, Takada K, Ito M, Nakajima N, Takahashi K, Lopes TJS, Dutta J, Khan Z, Kriti D, van Bakel H, Tokita A, Hagiwara H, Izumida N,Kuroki H,Nishino T,Wada N,Koga M,Adachi E,Jubishi D,木谷H,Kawaoka Y.流感A的变体降低了对日本患者分离的Baloxavir敏感性的变体,并通过呼吸道液滴进行拟合。NAT微生物5:27-33。 Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza VirusNAT微生物5:27-33。Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza Virus