XiaoMi-AI文件搜索系统
World File Search SystemAcinetobacter

根际杆菌杆菌的生物活性与抗生素抗生素抗生素抗生素
这项研究的重点是从巴格达市的根际土壤中分离出的鲍曼尼杆菌产生和纯化的铁载体,并与所选抗生素进行独立和结合评估其生物活性。使用Chrom琼脂,生化和生理测试进行细菌鉴定,并通过PCR扩增16S rDNA管家基因确认。在培养琥珀酸酯肉汤中的细菌后,使用乙酸乙酯提取铁载体,并通过HPLC纯化,在403 nm的波长下检测到。从下呼吸道感染中获得了总共38种细菌分离株,包括大肠杆菌,肺炎克雷伯氏菌,铜绿假单胞菌,铜绿杆菌,baumanniii,金黄色葡萄球菌,金黄色葡萄球菌和塞拉蒂亚和srratia marcesencens。用13种抗生素进行的抗生素敏感性测试显示,氨苄西林(65.7%)和头孢曲松(63.1%)的抗性率最高,而使用amikacin(15.7%)观察到最低的耐药性。对铁载体的协同活性与头孢曲松,头孢嗪和庆大霉素相结合,以针对多剂量抗性(MDR)分离株进行了测试。通过铁载体和庆大霉素与金黄色葡萄球菌的结合观察到了最显着的抗菌活性,而对鲍曼尼曲霉的效果最小。总之,从下呼吸道感染中成功鉴定出38种细菌分离株。铁酚与庆大霉素的结合表现出对金黄色葡萄球菌的显着抗菌活性,但对鲍曼尼曲霉的作用无效。

在Soetomo医院烧伤单位的烧伤患者中新出现的多药抗杆菌baumannii的危险因素
摘要简介:由于烧伤损伤本身的性质,包括长时间住院,使用抗生素,治疗程序等,烧伤损伤患者处于感染的高风险。在这个时代,鲍曼尼(Baumannii)(A.BA)引起的医院感染已显着增加。进行了这项研究,以研究微生物模式和烧伤多药耐药性(MDR)baumannii(A.BA)的燃烧患者的危险因素(A.BA)。材料和方法:我们从2020年1月至2021年12月在Soetomo医院的烧伤单位进行了回顾性观察性研究。通过单变量和多变量分析分析了MDR- A.BA的潜在危险因素。病例组包括诊断为MDR-A.BA伤口感染的患者。被诊断为非MDR的患者是:(1)患者除A.BA以外的其他微生物,(2)无菌分离株,以及(3)对照组中分离为A.BA但不包括MDR的患者。结果:这项研究总共包括120名烧伤患者。在这项研究中,发现有24%的烧伤患者患有鲍曼尼杆菌和79%(来自24%的baumannii的24%)患有MDR-A.BA。根据单变量分析,显着的危险因素是:缩写的烧伤严重程度指数(absi)(p = 0,002; or:6.10; CI:1,68-21,57);住院时间(LOS)(P <0,000; OR:6.95; CI:2,56-18,91)和合并症(P = 0,006; OR:3,72; CI:1,44-9,58)。关键字:烧伤,鲍曼尼杆菌,多药电阻但是,在通过多变量分析进行分析之后,仅abs是重要的因素(p = 0,010; or:1,70; CI:1,23-2,36)。结论:基于单变量分析,MDR-A.BA的重要危险因素为:absi,住院时间和合并期限。,但是在通过多元分析调整后,仅abs是重要的因素。

无疤痕去除大型抗性岛可导致抗生素的敏感性和增加的鲍曼尼杆菌的自然转化性
摘要在基因组成方面具有巨大的多样性,包括多种推定的抗生素耐药性基因,阿巴岛是鲍曼尼杆菌杆菌多药的潜在贡献者。但是,ABAR对抗生素耐药性和细菌生理学的有效贡献仍然难以捉摸。为了解决这个问题,我们试图准确删除Abar Islands并恢复其插入站点的完整性。为此,我们设计了一种多功能无疤的基因组编辑策略。我们在最近的两个鲍曼尼菌临床菌株中形成了这种遗传修饰:分别携带19.7 kbp和86.2 kbp的Abar1和Abar1岛的菌株AB5075和菌株AYE。然后,在父母菌株及其固定衍生物之间进行抗生素敏感性。通过该岛的开放阅读框(ORF)的预测功能所预期的,抗抗性的抗抗药性在野生型和ABAR11固定的AB5075菌株之间相同。ABAR1具有25个ORF,预测抗生素类别具有抗性,并且AYE ABAR1固定衍生物显示出对多种类别的抗生素的可疑性。此外,ABARS的固化恢复了高水平的自然转化性。的确,大多数阿巴群岛都被插入与自然转化有关的通讯基因中。我们的数据表明,Abar插入有效地失活,并且还原的通信是功能性的。固化始终导致高度转换,因此很容易遗传诱因。ABAR的修改提供了对Abar获取功能的洞察力的见解。

baumannii菌株的表型和分子检测产生脱氧教学医院临床标本的碳青霉酶
如今,细菌中的抗生素耐药性已成为一个全球问题。 因此,在选择更有效的治疗溶液中,鉴定细菌菌株引起了特别的关注。 抗药性最常见的机制之一是鲍曼尼杆菌杆菌酶的产生。 本研究旨在通过表型和分子方法检测碳纤维烯酶产生菌株,用于2021年6月至2022年6月之间在Dezful的Ganjavian医院收集的临床标本中。。如今,细菌中的抗生素耐药性已成为一个全球问题。因此,在选择更有效的治疗溶液中,鉴定细菌菌株引起了特别的关注。抗药性最常见的机制之一是鲍曼尼杆菌杆菌酶的产生。本研究旨在通过表型和分子方法检测碳纤维烯酶产生菌株,用于2021年6月至2022年6月之间在Dezful的Ganjavian医院收集的临床标本中。timicrobial易感性测试,而使用CEFTAZIDIME和CEFTAZIDIME /CLAVAVAZIPIMIMIMIMIMIC ADIPEN和IMIPENIP和IMIPSICEN和IMIPEN IMIIPEN和IMIPEN IMIPEN和IMIPEN IMIIPEN和IMIPEN IMIIPEN和IMIPEN IMIPCEN和IMIPEN,将扩展的β-内酰胺酶(ESBLS)和金属近似群(MBLS)进行了延长的谱。 分别。BLA IMP,BLA SPM,BLA OXA-23和BLA OXA-24,BLA OXA-58的分子检测进行了Bla oxa-58。总共54个菌株,与米诺环素相比(13%)相比,头孢菌素的最高电阻率为头孢菌素(98.1%)和环氧菌(94.2%)(94.2%)。ESBL和MBL生产者分别为26%和80%。所有分离株都对结肠菌素具有中间抗性。抗碳青霉烯曲霉(CRAB)中最普遍的基因是BLA OXA-23,其次是BLA AOXA-24,BLA GES,BLA GES,BLA IMP和BLA OXA-58基因。本报告强调了螃蟹和对结菌素的中间抗性的存在,以及该地区不同碳酸碳纤维酶类别的几个基因的共存。因此,应及时确定抗性菌株,并应设计特定的治疗方案以控制治疗环境中抗药性基因的传播。


沙特实验室专家许可检查(SLLE) div>
Acinetobacter, Aeromonas, Bordetella, Brucella , Campylobacter, Eikenella, Francisella, Haemophilus, Helicobacter, Legionella, Pasteurella, Plesiomonas, Pseudomonas, Burkholderia, Stenotrophomonas, Chryseobacterium, Vibrio, HACEK, Bartonella, capnocytophaga)

Brock-细菌中的遗传交换
•生理转化发生在自然界中,其中包括:图表:显示由于版权限制而移除的细菌组之间所证明的遗传互连。•1)链球菌•2)葡萄球菌•3)芽孢杆菌•4)ACINETOBACTER•5)血友病•6)Neisseria

五年研究
结果:在气管切开术之前,在42%的患者中检测到定植。最常见的微生物是鲍曼尼杆菌(40.5%),铜绿假单胞菌(19%)等(40.5%)。总共确定了184个病原体。定殖,8例患者中有3例病原体,43例患者的2例病原体和72例患者的单个病原体。此外,从气管弹式培养物中分离出13种不同的微生物,鲍曼尼(Baumannii)(38.6%),铜绿假单胞菌(18.5%)(18.5%)和serratia marcescens(8.1%)是最常见的,而其他微型机器人占34.8%。定殖患者的总体死亡率为60.5%,在内托氏气管抽吸培养物中检测到的鲍曼尼氏杆菌杆菌的死亡率最高(41.2%)。该组的缺血性或出血性脑损伤的发生率也更高(62.3%)。

Accugen检测常见的临床致病微生物
Acinetobacter Baumannii, Staphylococcus capnocytophaga Haemolytica, Pseudomonas fluorescens, Staphylococcus horses, Actinomyces Israelii, Staphylococcus Epidermidis, Capnocytophaga Ochracea, Pseudomonas Mosselii, Streptobacillus moniliformis, Bordetella tunnels,葡萄球菌血液溶血,囊孢子虫,pseudomonas putida,链球菌,Gallolyticus,Burkholderia cepacia,葡萄球菌,弯曲球菌,弯曲球菌Ococcus沙门氏菌肠道SSP。 div>Acinetobacter Baumannii, Staphylococcus capnocytophaga Haemolytica, Pseudomonas fluorescens, Staphylococcus horses, Actinomyces Israelii, Staphylococcus Epidermidis, Capnocytophaga Ochracea, Pseudomonas Mosselii, Streptobacillus moniliformis, Bordetella tunnels,葡萄球菌血液溶血,囊孢子虫,pseudomonas putida,链球菌,Gallolyticus,Burkholderia cepacia,葡萄球菌,弯曲球菌,弯曲球菌Ococcus沙门氏菌肠道SSP。 div>
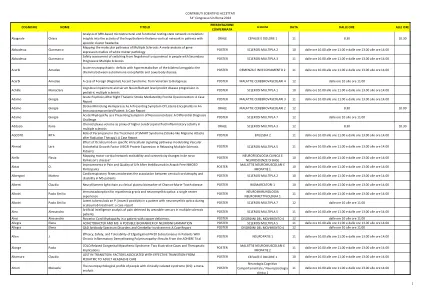
接受科学贡献第54届国会罪...
Alesi Alessandro后脐带骨髓病患有缺乏铜的患者。POSTER SPARTINI of Movement 4 12 from 10 to 11.00 Allegra Elena Acinetobacter and MS: A Possible Biomarker of NeurOinflammation Poster Multiple Sclerosis 5 11 from 10.00 to 11.00 and from 13.00 to 2.00 pm Allagra Elena Gad Antibody-Spectrum Disorders and Cerebelar Involvement: A Case Report Postter Postter of the Movement 4 12 from 10 to 11.00POSTER SPARTINI of Movement 4 12 from 10 to 11.00 Allegra Elena Acinetobacter and MS: A Possible Biomarker of NeurOinflammation Poster Multiple Sclerosis 5 11 from 10.00 to 11.00 and from 13.00 to 2.00 pm Allagra Elena Gad Antibody-Spectrum Disorders and Cerebelar Involvement: A Case Report Postter Postter of the Movement 4 12 from 10 to 11.00