XiaoMi-AI文件搜索系统
World File Search SystemAxon


开发基于细胞的测定方法,用于测量多酚处理细胞的NAD水平
青光眼是世界上视力丧失的主要原因之一,其特征是视网膜神经节细胞(RGC)的功能障碍。青光眼的早期病理机理是RGC的轴突的变性,发现可以预防轴突变性的新疗法引起了极大的关注。在许多神经退行性系统中,增加辅酶烟酰胺腺苷二核苷酸(NAD)的浓度已被证明是轴突保护性。增加NAD可以通过增加参与神经元NAD,烟酰胺单核苷酸腺苷转移酶2(NMNAT2)的末端酶的催化特性来实现。nmnat2是理想的治疗靶标。多酚A(PA),这是一种不会披露的多酚,已被证明是通过NMNAT2的正调制来提高NAD的。的目的是开发一种基于细胞的测定法,用于筛选PA和12个新型PA的类似物,以在从C57BL/6J小鼠中分离出脑皮质,视网膜和肝细胞中其NAD促进作用。进行了使用生物发光测定的方案,以优化变量,例如细胞浓度,底物(烟酰胺)浓度,PA浓度和孵育时间。该方法开发产生了一日测试PA及其在皮质细胞中类似物的方案。pa及其几个类似物表现出NAD促进效应。该方案以及筛选的结果可以进一步用于开发可预防青光眼以及其他轴突和神经退行性的新型药物。

e2424232024.full.pdf
无菌α和包含1(SARM1)的TIR基序是一种可诱导的NADASE,在损伤后发生的整个神经元和感觉代谢变化都定位于线粒体。在SARM1耗竭或激活后观察到最小的蛋白质组学变化,这表明SARM1不会对神经元蛋白稳态产生广泛的影响。然而,响应损伤和细胞胁迫的整个神经元中是否发生SARM1激活,在很大程度上未知。使用半小动的成像管道和定制的深度学习评分算法,我们研究了混合性性小鼠原发性皮质神经元和男性人类诱导的多能干细胞衍生的皮质神经元的变性,以响应许多不同的胁迫。我们表明,根据压力源,SARM1激活差异限于特定的神经元室。皮质神经元在机械横切后经历SARM1依赖性轴突变性,而SARM1激活仅限于损伤部位远端的轴突室。然而,VACOR处理后的全局SARM1激活会导致细胞体和轴突变性。上下文特异性应激源,例如微管功能障碍和线粒体应力,会诱导轴突SARM1激活,从而导致SARM1依赖性轴突变性和与SARM1无关的细胞体死亡。我们的数据表明,隔室特异性SARM1 - 介导的死亡信号传导取决于损伤的类型和细胞应激源。
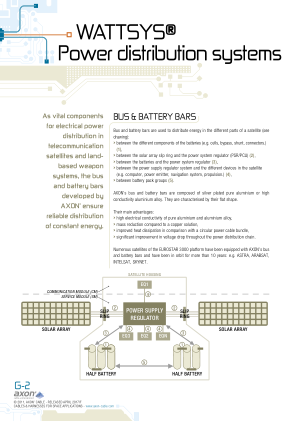
WATTSYS® 配电系统
为了将母线安装到航天器壁上,需要使用刚性铝制固定装置和由高性能热塑性聚合物或铝组件制成的柔性固定装置。这些有助于显著减少振动、冲击和温度的机械影响。为了安装母线,AXON' 建议每隔 200 毫米 (7.9”) 放置一个刚性固定装置和多个柔性固定装置。柔性固定装置允许母线随着航天器的运动而弯曲,同时考虑到其热弹性行为。刚性支撑确保母线完全固定到机械结构上。固定装置和母线的安装根据适用的客户规范进行。

在cat鱼的胸脑外侧皮质区域中多极神经元的形态状态,clarrias batrachus:一项高尔基研究
1。引言神经元是高度极化的细胞类型,在结构和功能上具有不同的过程,并从介导信息流过神经系统(例如树突和轴突)的SOMA延伸。轴突是一个类似线的过程,它通过从SOMA出现的神经递质的释放传输到其他神经元,这是一个单个长过程。来自Soma出现的多个分支过程称为树突。树突中包含神经递质受体,可从相邻连接的神经元收集信号[1]。神经元,其中三个以上的树突由soma产生,并以不同角度或不同的杆子辐射为多极神经元,其轴突末端包含多型突触囊泡[2],一种突触特征,一种突触特征,通常与抑制性神经转相者相关。在哺乳动物中,在锥体神经元之后,第二个位置由多极神经元获得[4]。

MYCBP2 的功能丧失变异导致神经行为表型和胼胝体缺陷
胼胝体是连接大脑两个半球的一束轴突纤维。以胼胝体发育不良为核心表型的神经发育障碍为了解轴突发育异常导致的病理学提供了宝贵的窗口。本文描述了一组八名患有神经发育障碍的患者,这些患者的特点是一系列缺陷,包括胼胝体异常、发育迟缓、智力障碍、癫痫和自闭症特征。每位患者都携带 MYCBP2 的一个独特的新生变异,MYCBP2 基因编码一种非典型 RING 泛素连接酶和信号传导中枢,在轴突发育中具有进化保守的功能。我们利用 CRISPR/Cas9 基因编辑将疾病相关变异引入秀丽隐杆线虫 MYCBP2 直系同源物 RPM-1 的保守残基中,并评估了体内功能结果。与 MYCBP2 变异患者的不同表型一致,携带 rpm-1 中相应人类突变的秀丽隐杆线虫表现出轴突和行为异常,包括习惯改变。此外,影响 RPM-1 泛素连接酶活性的变异中发生了自噬标记物 LGG-1/LC3 的异常轴突积累。解剖学、细胞生物学和行为读数的功能遗传结果表明,MYCBP2 变异可能会导致功能丧失。总的来说,我们从多名人类患者和在体动物模型上进行 CRISPR 基因编辑的结果支持了 MYCBP2 与人类神经发育谱系障碍之间的直接联系,我们称之为 MYCBP2 相关发育迟缓伴有胼胝体缺陷 (MDCD)。

寻找轴突的起源
锥体神经元很容易辨认,因为它们的胞体(神经元中包含细胞核的部分)具有特征性的三角形(因此得名)。然而,仔细检查就会发现,胞体的大小会有所不同,向胞体传递信号的树突所形成的“树突”的大小和形状也会有所不同(DeFelipe and Fariñas,1992 年)。此外,据报道,哺乳动物皮层中的一些锥体神经元的轴突从树突而不是胞体底部出现(Triarhou,2014 年;图 1)。这些“携带轴突的树突”很不寻常,因为树突接收的信号通常在胞体中经过处理,然后通过轴突发送到其他神经元(Förster,2014 年)。这些形态差异很重要,因为它们会影响单个神经元和神经元组计算信息的方式。研究人员对只发生在人类和灵长类动物身上的特征特别感兴趣,因为这些特征可能与认知有关

将少突胶质细胞的功能扩展到大脑能量代谢
要紧密地拉开(图1 C)。这允许众所周知的盐酸在兰维尔的一个节点到下一个节点的作用电位传播,轴突离子电流还涉及少突胶质细胞中的胞质细胞质空间[2]。轴突髓鞘形成加快神经传导速度的速度与轴突直径的函数[3],因此,髓磷脂特别与大脊椎动物的演化特别相关,这些大脊椎动物需要在长距离远距离进行快速的轴突传导。脊椎动物髓磷脂发生在系统发育时间轴中相对较晚,但是在几乎所有其他门的门中都观察到了轴突的神经胶片[4]。轴突覆盖神经胶质细胞也被认为可以防止相邻轴突之间的“边缘耦合”,即通过密切接触未定期激活。在脊椎动物中,这可能与并行[5](例如视神经,callosum callosum或脊髓的大型轴突段)有关,但是对此假设的实验支持很难获得。

轴突张力有助于一致的折叠放置
尽管存在轴突行为的证据,但实验结果对轴突张力假说的全面接受提出了挑战。例如,在成年小鼠 18 和发育中的雪貂 19 的大脑中进行的残余应力切割实验表明,皮层下存在持续的张力,这可能对折叠过程产生重大影响。然而,三个主要结论挑战了基于张力的折叠假说:(1)皮层下轴突张力远离折叠区域,(2)脑回周围的周向轴突张力太弱,无法直接拉动组织,(3)观察到的脑回中残余应力的方向与模型的预测不符。19 他们的实验和模拟表明差异生长主要驱动折叠,同时允许轴突张力仍然是影响皮质折叠的制约因素。在其他研究中,轴突连接被发现与跨物种的皮质折叠成比例,20,21 导致研究人员扩展原始的轴突张力理论,提出轴突张力导致白质折叠,进而影响灰质折叠。最近,Van Essen 重新表述了原始的基于张力的形态发生理论,在细胞和组织尺度上纳入了更多促进折叠的力量。22 反驳对其理论的批评,19 他指出,体外实验可能无法捕捉体内张力,这可能会受到切片或组织水肿的影响。他还呼吁建立一个模拟框架,能够模拟皮质组织中的关键神经生物学特征,例如以不同角度甚至交叉取向的轴突。23 目前,在理解轴突张力在脑回形成过程中如何发挥作用方面仍然存在差距。例如,体内存在什么程度的轴突张力?这种张力水平是否能够触发皮质折叠?轴突网络在折叠过程中如何连接?鉴于有关大脑结构和功能之间关系的悬而未决的问题,白质尤其令人感兴趣。24 据观察,各种神经系统疾病中都存在异常的白质连接,这通常与大脑内的非典型折叠模式相吻合。当然,这些关系可能是因果关系,也可能仅仅是相关的。无论如何,更深入地了解白质连接在皮质折叠中的作用,对我们理解大脑的结构和功能具有深远的影响。
