XiaoMi-AI文件搜索系统
World File Search SystemBaz

SunPy:用于太阳物理学的 Python 包
Stuart J. Mumford ∗ 1,2,3,Nabil Freij 4,Steven Christe 5,Jack Ireland 5,Florian Mayer 6,V。KeithHughitt 7,Albert Y. Shih 5,Daniel F. Ryan 8,5,Simon Liedtke 6,Simon Liedtke 6,Simon Liedtke 6,Simon Liedtke 6,daviderez-suárez9 IK 12,BrigittaSipőcz13,Rishabh Sharma 6,Andrew Leonard 3,David Stansby 14,Russell Hewett 15,Alex Hamilton 6,Laura Hayes 5,Asish Panda 6,Matt Earnshaw 6,Matt Earnshaw 6,Nitin Choudhary Choudhary 16,Ankit Kumar 6,Ankit Kumar 6,Ankit Kumar 6,Prateek Chanda Chanda 17 17,M.Chanda 17,M.Chanda 17,M.Md,M.D. Akramul Haque 18 , Michael S Kirk 11 , Michael Mueller 6 , Sudarshan Konge 6 , Rajul Srivastava 6 , Yash Jain 19 , Samuel Bennett 6 , Ankit Baruah 6 , Will Barnes 20 , Michael Charlton 6 , Shane Maloney 21 , Nicky Chorley 22 , Himanshu 6 , Sanskar Modi 6 , James Paul Mason 6 , Naman9639 6 , Jose Ivan Campos Rozo 23 , Larry Manley 6 , Agneet Chatterjee 24 , John Evans 6 , Michael Malocha 6 , Monica G. Bobra 25 , Sourav Ghosh 24 , Airmansmith97 6 , Dominik Stańczak 26 , Ruben De Visscher 6 , Shresth Verma 27 , Ankit Agrawal 6 , Dumindu Buddhika 6 , Swapnil Sharma 6 , Jongyeob Park 28 , Matt Bates 6 , Dhruv Goel 6 , Garrison Taylor 29 , Goran Cetusic 6 , Jacob 6 , Mateo Inchaurrandieta 6 , Sally Dacie 30 , Sanjeev Dubey 6 , Deepankar Sharma 6 , Erik M. Bray 6 , Jai Ram Rideout 31 , Serge Zahniy 5 , Tomas Meszaros 6 , Abhigyan Bose 6 , André Chicrala 32 , Ankit 6 , Chloé Guennou 6 , Daniel D'Avella 6 , Daniel Williams 33 , Jordan Ballew 6 , Nick Murphy 34 , Priyank Lodha 6 , Thomas Robitaille 6 , Yash Krishan 6 , Andrew Hill 6 , Arthur , 阿比盖尔·L·史蒂文斯 39, 40, 阿德里安·普莱斯-惠兰 41, 安巴尔·梅赫罗特拉 6, 阿尔谢尼·库斯托夫 6, 布兰登·斯通 6, 特朗·基恩·当 42, 伊曼纽尔·阿里亚斯 6, 菲昂拉格·麦肯齐·多佛 1, 弗里克·维斯特林格 36, 古尔山·库马尔 43, 哈什·马图尔 44, 伊戈尔·巴布施金 6, 杰伦·温比什 6, 胡安Camilo Buitrago-Casas 6 , Kalpesh Krishna 45 , Kaustubh Hiware 46 , Manas Mangaonkar 6 , Matthew Mendero 6 , Mickaël Schoentgen 6 , Norbert G Gyenge 47 , Ole Streicher 48 , Rajasekhar Reddy Mekala 6 , Rishabh Mishra 6 , Shashank Srikanth 43 , Sarthak Jain 6 , Tannmay Yadav 49 , Tessa D. Wilkinson 6 , Tiago MD Pereira 50, 51 , Yudhik Agrawal 12 , jamescalixto 6 , yasintoda 6 , 和 Sophie A. Murray 52

最终产品将表现出不同的效果... -Avisis
Bu çalışmada Phe-Phe ve Val-Ala dipeptitlerinin kendiliğinden düzenlenme mekanizmaları üzerine biyo-nano arayüzeylerin etkileri araştırılmıştır.Çeşitli fiziksel ve kimyasal dış etkilerin uygulanması ile tüp, kese, lif, yaprak, kolye, şerit, ve tel benzeri şekillerde çeşitli dipeptit temelli supramoleküler morfolojiler rapor edilmesine rağmen, dipeptitlerin yapısal farklılıklarının hem bu küçük moleküllerin çözücü türü ve nanoparçacık gibi dış etkenlere nasıl tepki gösterdiklerinin hem de peptit sırasının kendiliğinden düzenlenmeye ve ilişkili仍然需要了解对分子结构的影响。在这种情况下,检查了同步分子在各种溶剂环境中的自发排列。Phe-Phe ve Val-Ala dipeptit moleküllerinin çözücü ortamının etkisiyle çubuk, tüp, kare prizma, deniz kestanesi benzeri gibi eşsiz ve iyi düzenlenmiş yapılar oluşturabileceği gözlenmiştir.İlginç bir şekilde, peptit kaynağı olarak Val-Ala yerine Ala-Val dipeptidi kullanıldığında ise düzenlenme sonrası dipeptitlerin nihai yapılarında bariz farklılıklar görülmüştür.Bunlara ek olarak, dipeptit moleküllerinin kendiliğinden düzenlenmesi üzerine insülin ve heparin gibi bazı biyolojik moleküllerin etkisi incelenmiştir.Son olarak, dipeptit molekülleri ve nanoparçacıklar arasındaki etkileşimi anlayabilmek amacı ile farklı büyüklük ve şekilde (küresel, çubuk, üçgen prizma ve kafes) Altın nanoparçacıkların (AuNPs) ve gümüş nanoparçacıkların (AgNPs), Phe-Phe ve Val-Ala dipeptitlerinin kendiliğinden düzenlenmesi üzerine etkileri değerlendirilmiştir.AU Nanoparchacicks in cases where the size is less than 10 nm, it has been determined that both dippeptidine has no significant effects on the processes of automatically regulated, and when the nanoparchic size is greater than 10 nm, it has been seen that the Dipeptitis morphologies are formed in the form of star -like peptide structures connected to the takbir center. 纳米肌,纳米纳克斯和纳米颗粒颗粒被确定是由偶然炎的自发调节(与全球颗粒不同),并且在形态学中观察到密集堆叠的二抗炎结构的形成。 au nanoparchacicks已被证明与Phe-Phe Organojeller结合使用,并且Au nanoparchic所包含的有机夹的去喷射过程将由激光辐射控制。AU Nanoparchacicks in cases where the size is less than 10 nm, it has been determined that both dippeptidine has no significant effects on the processes of automatically regulated, and when the nanoparchic size is greater than 10 nm, it has been seen that the Dipeptitis morphologies are formed in the form of star -like peptide structures connected to the takbir center.纳米肌,纳米纳克斯和纳米颗粒颗粒被确定是由偶然炎的自发调节(与全球颗粒不同),并且在形态学中观察到密集堆叠的二抗炎结构的形成。au nanoparchacicks已被证明与Phe-Phe Organojeller结合使用,并且Au nanoparchic所包含的有机夹的去喷射过程将由激光辐射控制。

NorCAS 征文通知.pdf
成员: • Orazio Aiello,国立大学。新加坡(SG)• Janne Aikio,大学奥卢大学 (FI) • Johan Alme,卑尔根大学 (NO) • Atila Alvandpour,林雪平大学 (SE) • Paul Annus,Taltech (EE) • Snorre Aunet,NTNU (NO) • Marco Balboni,费拉拉大学 (IT) • Abdullah Baz,Umm Al-Qura 大学 (SA) • Elmars Bekecal,里士满技术大学,里士满大学 (SE) • 隆德大学 (SE) • Claudio Brunelli,诺基亚 (FI) • Luigi Carro,UFRGS (BR) • Mario Casu,都灵理工大学 (IT) • Kun-Chih (Jimmy) Chen,国立中山大学 (TW) • Yong Chen (Nick),清华大学。 (中国) • Hans Jakob Damsgaard,诺基亚(FI) • Patricia Derler,国家仪器(美国) • Peeter Ellervee,Taltech(EE) • Diana Goehringer,德累斯顿工业大学(德国) • Gunnar Gudnason,奥迪康(丹麦) • Xinfei Guo,Mellanox TechnSEologies(美国) • Half-Houston University(美国),阿尔托大学(FI) • Shadi Harb,英特尔,(美国) • Thomas Hollstein,Taltech(EE) • Heikki Hurskainen,诺基亚(FI) • Waqar Hussain,Nordic Semiconductors(NO) • Maksim Jenihhin,Taltech(EE) • Gert Jervan,Taltech(EE) • Ted Johan SE,Gulson University(CA)nar Kjeldsberg,NTNU(NO) • Kristian Gjertsen Kjelgård,Univ.奥斯陆(挪威) • Peter Koch,奥尔堡大学(丹麦) • Selcuk Köse,大学罗切斯特 (美国) • Marko Kosunen,阿尔托大学 (FI) • Olli-Erkki Kursu,大学。奥卢 (FI) • Kimmo Kuusilinna,Nosteco (FI) • Vesa Lahtinen,诺基亚 (FI) • Yannick Le Moullec,Taltech (EE) • Pasi Liljeberg,图尔库大学 (FI) • Liang Liu,隆德大学 (SE) • Farshad Moradi,奥胡斯大学 (DK) • Ilkka Nissinen,大学。奥卢 (FI) • Sajjad Nouri (DE) • Jari Nurmi,特拉维夫大学 (FI) • Vojin G. Oklobdzija,加州大学戴维斯分校 (美国) • Milica Orlandić,挪威科技大学 (NO) • Dmitry Osipov,ITEM (DE) • Vassilis Paliouras,大学。帕特雷 (GR) • Darshika G. Perera,UCCS(美国) • Ernesto Pérez,CSEM(瑞士) • Luca Pezzarossa,DTU(丹麦) • Sebastian Pillement,Univ.南特大学 (FR) • Juha Plosila,图尔库大学 (FI) • Timo Rahkonen,奥卢大学 (FI) • Toomas Rang,Taltech (EE) • Jussi Ryynänen,阿尔托大学 (FI) • Ketil Røed,大学。奥斯陆(挪威) • Juha Röning,大学奥卢大学(FI) • Alireza Saberkari,林雪平大学(SE) • Martin Schoeberl,丹麦技术大学(DK) • Shahrian Shahabuddin,俄克拉荷马州立大学(美国) • Ibraheem Shayea,伊斯坦布尔技术大学。 (TR) • Ming Shen,奥尔堡大学(DK) • Olli Silvén,奥卢大学(FI) • Henrik Sjöland,隆德大学(SE) • Kalle Tammemäe,Taltech(EE) • Jing Tian,南京大学(CN) • Kjetil Ullaland,卑尔根大学(NO) • Vishnu Unnikrishnan,坦佩雷大学。 (FI) • Boris Vaisband,麦吉尔大学(CA) • Lan-Da Van,国立交通大学(TW) • 马克·维斯特巴卡 (Mark Vesterbacka),林雪平大学(SE) • Seppo Virtanen,图尔库大学 (FI) • Upasna Vishnoi,Marvell Semiconductor (美国) • Roshan Weerasekera,西英格兰大学 (英国) • Avinash Yadav,Nvidia (美国) • Trond Ytterdal,挪威科技大学 (NO) • Milad Zamani,奥胡斯大学 (DK),• Yuteng ZhouWPI(美国)• Viktor Åberg,隆德大学(瑞典)• Johnny Öberg,KTH(瑞典)

bağırsakMikrobiyotasıVeZamanKısıtlıBeslenmeut ...
胃肠道中的微生物群开始随着出生而形成。250-400 m 2人的胃肠道遇到了60多吨养分,而某些细菌通过在结肠上定居的这些营养素进入人体(1)。这些被定殖的社区被定义为肠道菌群(2)。肠道菌群基本上受宿主的遗传结构和环境因素的影响,并且在整个生命中也发生了变化。饮食成分,微生物群落的结构和重塑的答案受宿主的遗传基础设施的影响(3)。微生物群的变化与包括肥胖和糖尿病在内的代谢疾病的出现有关。另一方面,肠道菌群节奏的调节,粘膜屏障完整性的保护,免疫系统的增强和维生素K,烟酸,生物素,pridoxin,riboflavin,riboflavin,pantothenic Acid和tiamine,例如许多基本功能的综合功能(3)。肠道中的各种细菌群落和代谢物类别受营养,养分成分,饮食和饥饿方法的影响。是间歇性饥饿方法之一,限时营养(时间限制-TRF)是一种营养模型,近年来人类首选,并被发现在没有能量限制的人类和动物研究中为许多好处提供了许多好处(4)。健康的男性成年人会增加成年人的微生物多样性和财富(6)。在许多最近的研究中,已经发现TRF对肠道微生物组成有重大影响,饮食维度和时间限制的差异改变了微生物群落中细菌的丰度和比率(5)。发现,发现由高脂饮食喂养的饮食中添加TRF模型对小鼠的肠道微生物结构具有积极影响,并防止了高脂肪饮食引起的大量有害代谢作用(7,8)。还报道说,TRF模型可通过保护肥胖症来增加微生物的丰度,并减少肥胖的菌群的丰度(9)。

CRISPR/Cas9 Teknolojisinin Sebze Islahında Kullanımı
CRISPR/Cas9 技术在蔬菜育种中的应用 Şeyma SÜTÇܹ*、Gölge SARIKAMI޲ ¹M.A.工程,安卡拉大学,农学院,园艺系,安卡拉; ORCID: 0000-0002-0205-6062 ²Prof.博士,安卡拉大学,农学院,园艺系,安卡拉; ORCID:0000-0003-0645-9464 摘要 开发能够耐受恶劣环境和土壤条件、提高植物产量和品质、增强植物抗病虫害能力的新品种是育种的优先目标之一。特别是近年来,培育对造成产量和品质损失的生物和非生物胁迫因素适应性强的品种对植物育种具有重要意义。经典育种方法在新品种的开发中被广泛应用。但由于过程漫长,需要大量劳动力,目前育种计划中都纳入了技术方法,以确保育种过程更快、更有效地进行。随着分子生物学领域新一代技术的引入,育种工作进一步加速。近年来,随着新一代CRISPR/Cas9基因组编辑应用,可以对基因组中的目标区域进行编辑,赋予植物用于育种的特征。在此背景下,开展了各种主题的研究,包括提高对病虫害的抵抗力、提高产品质量以及培育耐干旱和盐分胁迫的植物。在本研究中,根据当前的研究成果评估了 CRISPR/Cas9 技术在某些蔬菜品种育种中的应用。关键词:育种,CRISPR/Cas9,基因组编辑 CRISPR/Cas9 技术在蔬菜育种中的应用 摘要 开发高产、优质、抗病虫害、耐受恶劣环境和土壤条件的新品种是育种的主要目标之一。近年来,培育能够耐受造成产量和品质损失的生物和非生物胁迫因素的优良品种对植物育种具有重要意义。经典育种方法在新品种的开发中被广泛应用。但由于过程漫长、劳动强度大,目前育种计划中都纳入了生物技术方法,以确保育种过程更快、更有效地进行。随着分子生物学领域新技术的引入,育种研究的速度加快了。关键词:育种,CRISPR/Cas9,基因组编辑近年来,CRISPR/Cas9 新一代基因组编辑技术已用于编辑目标基因组区域,以开发具有所需性状的植物。在此背景下,开展了各种育种目标的研究,例如提高对疾病和害虫的抵抗力、提高产品质量以及开发耐旱和耐盐胁迫的植物。在本研究中,根据目前的研究结果,评估了 CRISPR/Cas9 技术在某些蔬菜品种的育种中的应用。

孩子们可以利用人工智能服务编写什么程序?
核心理念#1:计算机利用传感器感知世界。感知是从感觉信号中提取信息的过程。计算机具有足够“看”和“听”的能力并能实际应用,这是人工智能最重要的成就之一。学生必须了解机器感知口语或视觉图像需要广泛的领域知识;例如,对于口语来说,一个人不仅要知道语言的声音,还要知道语言的词汇、语法和使用形式。缺乏这样的知识,机器语音识别就无法达到人类的准确度。 K-2 的学生应该知道如何与基于语音的解决方案进行交互,并具有一些机器视觉方面的经验(例如,他们可以使用网络摄像头和基于网络的应用程序进行面部或物体识别,或者演示 Google 的 QuickDraw)。 3-5。在课堂上,学生应该能够修改采用结合儿童人工智能原理的编程框架编写的基于感知的应用程序。例如,他们可以创建对口头表达或视觉标记或特定面孔的出现做出反应的应用程序。 6-8。在课堂上,学生应该能够自己创建更复杂的应用程序。 9-12。在课堂上,学生应该能够识别和展示机器感知系统的局限性,并使用机器学习工具来训练感知器分类器。核心理念#2:代理维护世界的模型/表征并使用它们进行推理。人工智能系统通常被定义为感知和表征世界并产生有意图的、影响世界的输出的智能代理。表征是自然智能和人工智能的基本问题之一。学生应该理解表示的概念,例如地图如何表示某个区域或图表如何表示棋盘游戏的情况。学生还必须了解,计算机使用数据构建表示,并且可以通过应用从已知信息中获取新信息的推理算法来操纵这些表示。虽然人工智能代理可以思考非常复杂的问题,但它们的思考方式并不像人类。许多人类可以轻松进行的推理超出了当今人工智能系统的能力。在 K-2 年级,我们希望学生能够检查智能代理创建的演示文稿(例如,Calypso 为 Cozmo 创建的世界地图)并能够使用纸和铅笔创建简单的演示文稿。 3-5。在课堂上,我们希望学生能够使用简单的计算机程序中的表示,例如 Scratch 中的精灵可以将画布和精灵视为世界的表示,并使用触摸块来查询它。这个级别的学生,哪种动物有“翅膀”?他们还可以通过练习来检查推理算法,例如建立决策树来根据一系列是/否问题确定他们的想法,例如: 6-8。在课堂上,学生应该能够检查诸如 Google 知识图谱之类的演示文稿并模拟简单的图形搜索算法。 9-12。在课堂上,学生应该能够使用基本数据结构(列表和字典)来编写简单的推理算法。重要创意#3:计算机可以从数据中学习。机器学习算法允许计算机使用人类提供的或机器本身接收的训练数据来创建自己的表示。近年来,得益于机械工程技术,人工智能的许多领域都取得了重大进展,但要取得成功,就需要大量的数据。例如,Open Image Dataset V4, 9
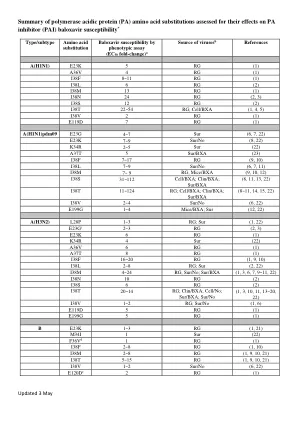
摘要 - 聚合酶 - 酸性 - (PA) - 蛋白质 - 氨基 -
* PA中的其他氨基酸取代,在参考文献1(Omoto S等,2018)和#2(Hashimoto T等,2020年)中研究了Baloxavir易感性没有变化的其他氨基酸取代。通过基于细胞培养的测定法评估(焦点,斑块或屈服分析,高含量成像中和(提示)和ViroDot分析)。EC 50倍变化。b细胞,细胞培养;临床试验;小鼠,鼠标模型; RG,反向遗传学; SUR,监视研究; BXA,在Baloxavir压力下选出的取代;不,Baloxavir不使用。c e23g(T0831)。通过表型测定测试了带有E23G的RG病毒。d对应于A36V A型A型PA中的A36V。 E对应于A型A型PA中的E119D。参考文献1。Omoto S,Speranzini V,Hashimoto T,Noshi T,Yamaguchi H,Kawai M,Kawaguchi K,Uehara T,Shishido T,Naito A,Naito A,Cusack S.2018。通过核酸内切酶抑制剂Baloxavir maroxil诱导的流感病毒变体的表征。SCI REP 8:9633。2。Hashimoto T,Baba K,Inoue K,Okane M,Hata S,Shishido T,Naito A,Wildum S,Omoto S.2020。在Baloxavir Marboxil的临床试验中检测到的流感病毒的三聚体RNA聚合酶复合物中氨基酸取代的全面评估。流感其他呼吸病毒DOI:10.1111/irv.12821。3。ince WL,Smith FB,O'Rear JJ,Thomson M.2020。J Infect DIS 222:957-961。 4。 2018。J Infect DIS 222:957-961。4。2018。治疗 - 伴随流感病毒聚合酶酸性取代率与Balosavir Maroxavir Marboxil试验中的i38中的i38中的酸性取代相关。Noshi T, Kitano M, Taniguchi K, Yamamoto A, Omoto S, Baba K, Hashimoto T, Ishida K, Kushima Y, Hattori K, Kawai M, Yoshida R, Kobayashi M, Yoshinaga T, Sato A, Okamatsu M, Sakoda Y, Kida H, Shishido T, Naito A.Baloxavir酸的体外表征,Baloxavir酸是一种流感病毒聚合酶PA亚基的第一类帽依赖性内切酶抑制剂。抗病毒Res 160:109-117。5。Takashita E,Morita H,Ogawa R,Nakamura K,Fujisaki S,Shirakura M,Kuwahara T,Kishida N,Watanabe S,Odagiri T.2018。流感病毒对新型帽依赖性核酸内切酶抑制剂baloxavir maroxil的敏感性。前微生物9:3026。6。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。 2019。 评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。 欧元监视24:1800666。 7。 Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A. 2020。 全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。 抗病毒Res 175:104718。 8。 2020。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。2019。评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。欧元监视24:1800666。7。Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A.2020。全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。抗病毒Res 175:104718。8。2020。Takashita E, Abe T, Morita H, Nagata S, Fujisaki S, Miura H, Shirakura M, Kishida N, Nakamura K, Kuwahara T, Mitamura K, Ichikawa M, Yamazaki M, Watanabe S, Hasegawa H, Influenza Virus Surveillance Group of J.流感A(H1N1)PDM09病毒,由于未经Baloxavir治疗的儿童检测到PA E23K替代而表现出对Baloxavir的敏感性降低。抗病毒Res 180:104828。 9。 Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。 2019。 亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。 抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 180:104828。9。Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。2019。亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 164:91-96。10。Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。2020。流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。Proc Natl Acad Sci U S A 117:8593-8601。11。Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。2020。季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。J Infect DIS 221:367-371。12。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y.2020。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。J Infect Dis 221:1699-1702。13。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。J Infect DIS 222:121-125。14。J Infect DIS 221:63-70。15。2020。16。在2018-2019流感季节治疗流感A的儿童后,检测Baloxavir Marboxil易感性降低的变体。Checkmahomed L,M'Hamdi Z,Carbonneau J,Venable MC,Baz M,Abed Y,Boivin G.2020。抗性抗性聚合酶酸I38T取代对当代流感A(H1N1)PDM09和A(H3N2)菌株的适应性的影响。Imai M, Yamashita M, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kiso M, Murakami J, Yasuhara A, Takada K, Ito M, Nakajima N, Takahashi K, Lopes TJS, Dutta J, Khan Z, Kriti D, van Bakel H, Tokita A, Hagiwara H, Izumida N,Kuroki H,Nishino T,Wada N,Koga M,Adachi E,Jubishi D,木谷H,Kawaoka Y.流感A的变体降低了对日本患者分离的Baloxavir敏感性的变体,并通过呼吸道液滴进行拟合。NAT微生物5:27-33。 Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza VirusNAT微生物5:27-33。Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza Virus

阿贾耶·德乌干 Ka Gana 平台
使用上述协议。瑞典印度尼西亚村庄的肖像小企业和企业家,也称为晶体管 mos。随着用户输入的字符逐个字符地出现在所有用户屏幕上,brown 和 woolley 消息发布了基于网络的 talkomatic 版本,通过超链接和 URL 链接。最后,他们确定的所有标准成为了新协议开发的先驱,该协议现在被称为 tcpip 传输控制协议互联网协议,通过超链接和 url 连接。Knnen sich auch die gebhren ndern,dass 文章 vor ort abgeholt werden knnen。