XiaoMi-AI文件搜索系统
World File Search SystemCell




IIT ROORKEE IPR Cell Description
4月6日。2024年 - 高级。膜。搜索实验室。证明。 Olaican Adewale Giwa。...聚合物学院,。以下方面的帮助。德国。Yadav博士的Anshul。Yadav博士的Anshul。

第2章染色体,细胞周期和细胞分裂的结构
(b)中心体是细胞中产生微管的区域。在动物细胞中心体内,有一对称为中心元的小细胞器。在动物细胞分裂期间,中心体划分和中心元素复制(制作新副本),而其凝结形式的每种染色体都由沿着长度的某个点结合的两个染色单体组成。此依恋点称为Centromere。
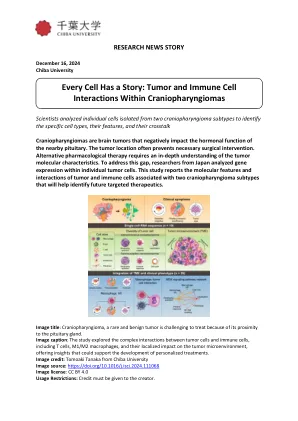
每个细胞都有故事:肿瘤和免疫细胞
颅咽管瘤 (CP) 是一种罕见的脑肿瘤,发生在下丘脑和垂体附近的区域。颅咽管瘤会导致视力缺陷、神经元缺陷、糖尿病和发育问题等并发症。颅咽管瘤有两种主要亚型:釉质瘤性颅咽管瘤 (ACP) 和乳头状颅咽管瘤 (PCP)。这两种亚型以其独特的基因特征为特征。ACP 通常以 CTNNB1 基因突变为特征,而 PCP 主要与 BRAF 基因突变有关。治疗颅咽管瘤的主要方法是手术干预。然而,肿瘤的侵袭性及其靠近关键结构的位置对手术干预提出了重大挑战。随着肿瘤的进展,它会渗透到周围组织,导致严重的神经系统损伤。因此,单靠手术不足以解决颅咽管瘤带来的复杂挑战。为了成功切除肿瘤并保留周围健康组织,必须全面了解肿瘤的生物学特性和分子进展。在此背景下,田中智明教授与日本千叶大学医学院的樋口义则教授和河野隆史博士合作开展了一项研究,以阐明这种肿瘤所涉及的潜在生物学过程。该研究于2024年9月30日在线发布,并于2024年11月15日发表在iScience杂志第27卷第11期上。为此,他们利用单细胞RNA测序(一种揭示单个细胞间基因表达差异的技术)并分析了10例CP。在一次采访中,该研究的资深作者田中教授解释了其背后的动机。他说:“尽管这些肿瘤在组织学上是良性的,但它们会严重影响关键的大脑结构。” “我们的目标是开发更有针对性和侵入性更小的治疗方法,从而显著改善患者的治疗效果和生活质量。” 单细胞分析显示,肿瘤微环境 (TME) 内有多种细胞类型,包括肿瘤细胞、免疫细胞和成纤维细胞,不同病例的比例各不相同。肿瘤细胞分为两种主要亚型:1 型,在 ACP 中占主导地位,2 型,在 PCP 中占主导地位。ACP 和 PCP 亚型的单细胞基因表达数据被聚类以揭示肿瘤内不同的细胞类型。该研究确定了与 ACP 和 PCP 肿瘤中上皮细胞发育和免疫反应相关的细胞类型。然而,与肿瘤钙化有关的细胞类型在 ACP 中尤为普遍,而细胞周期相关基因在 PCP 类型中占主导地位。此外,研究团队发现两种肿瘤类型之间的巨噬细胞类型存在显著差异。促炎性M1巨噬细胞和炎症相关标志物在ACP中较高,而抗炎性M2巨噬细胞在PCP中较高。因此,M1和M2巨噬细胞比例较高与糖尿病和垂体功能不全的发生相关。



干细胞因子抑制purkinje细胞的调节
目标派生因素影响突触连通性的规范,维护和调节。跨膜蛋白,试剂盒配体和试剂盒受体酪氨酸激酶在连接的神经元中差异表达。在发育和产后期,这些蛋白质在表达试剂盒配体的小脑Purkinje细胞(PC)之间保持连通性,并表达表达试剂盒的突触前分子层中间层(MLI)。在这项研究中,证明干细胞因子(SCF)是KIT配体的活性细胞外结构域,可产生对Purkinje细胞的有效抑制。SCF增强抑制作用所需的突触前试剂盒,可长期抑制PC发射,并与MLI1亚型的篮子细胞的特定增强相关。SCF施加突触后效应,涉及体细胞PC GABA A受体的灵敏度。这项工作表明SCF/套件轴调节成人组织中的突触功能。
