XiaoMi-AI文件搜索系统
World File Search SystemClpP

变构线粒体CLPP激动剂ONC206改变了压力反应,代谢和表观遗传学特征,以在高级GL
参考文献1。Allen Je和Al。SCI Transl Med。2013; 5(171):1717; 2。 rb冻结和al。 Pharmacol's。 2021; 100:372-387; 3。 ns疯狂和al。 nat公社。 2019; 10:5221; 4。 Ishizawa J和Al。 癌细胞。 2019; 35:721-737 E9; 5。 PR仪式和Al。 ACS头。 2019; 14:1020-1029; 6。 chi as和al。 j神经。 2019; 145(1):97-105; 7。 theeler bj和al。 J Clin Oncol.2020; 8。 vv prabhu和al。 歌手res。 2020; 80(16_supplementary):5688-5688; 9。 Wagner J和Al。 循环单元。 2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2013; 5(171):1717; 2。rb冻结和al。Pharmacol's。2021; 100:372-387; 3。ns疯狂和al。nat公社。2019; 10:5221; 4。 Ishizawa J和Al。 癌细胞。 2019; 35:721-737 E9; 5。 PR仪式和Al。 ACS头。 2019; 14:1020-1029; 6。 chi as和al。 j神经。 2019; 145(1):97-105; 7。 theeler bj和al。 J Clin Oncol.2020; 8。 vv prabhu和al。 歌手res。 2020; 80(16_supplementary):5688-5688; 9。 Wagner J和Al。 循环单元。 2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2019; 10:5221; 4。Ishizawa J和Al。癌细胞。2019; 35:721-737 E9; 5。 PR仪式和Al。 ACS头。 2019; 14:1020-1029; 6。 chi as和al。 j神经。 2019; 145(1):97-105; 7。 theeler bj和al。 J Clin Oncol.2020; 8。 vv prabhu和al。 歌手res。 2020; 80(16_supplementary):5688-5688; 9。 Wagner J和Al。 循环单元。 2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2019; 35:721-737 E9; 5。PR仪式和Al。ACS头。2019; 14:1020-1029; 6。 chi as和al。 j神经。 2019; 145(1):97-105; 7。 theeler bj和al。 J Clin Oncol.2020; 8。 vv prabhu和al。 歌手res。 2020; 80(16_supplementary):5688-5688; 9。 Wagner J和Al。 循环单元。 2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2019; 14:1020-1029; 6。chi as和al。j神经。2019; 145(1):97-105; 7。 theeler bj和al。 J Clin Oncol.2020; 8。 vv prabhu和al。 歌手res。 2020; 80(16_supplementary):5688-5688; 9。 Wagner J和Al。 循环单元。 2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2019; 145(1):97-105; 7。theeler bj和al。J Clin Oncol.2020; 8。vv prabhu和al。歌手res。2020; 80(16_supplementary):5688-5688; 9。Wagner J和Al。循环单元。2017; 16:1790-1799; 10。 Staley A和Al。 AM J Singing Res。 2021; 11(11):5374-5387; 11。 张Y和Al。 Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。2017; 16:1790-1799; 10。Staley A和Al。AM J Singing Res。2021; 11(11):5374-5387; 11。张Y和Al。Oncol Front。 2020; 10:57141; 12。 Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。Oncol Front。2020; 10:57141; 12。Tucker K和Al。 AM J Singing Res。 2022; 12(2):521-536; 13。 vv prabhu和al。 Clins Ress。 2019; 25:2305-2313; 14。 Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。Tucker K和Al。AM J Singing Res。2022; 12(2):521-536; 13。vv prabhu和al。Clins Ress。2019; 25:2305-2313; 14。Jassal B和Al。 九十res。 2020 JAN 8; 48(D1):D498-D503。Jassal B和Al。九十res。2020 JAN 8; 48(D1):D498-D503。2020 JAN 8; 48(D1):D498-D503。

线粒体基质蛋白酶 ClpP 激动剂通过破坏线粒体稳态来抑制乳腺癌细胞中的癌症干细胞功能
105 并且也可根据 CC0 许可使用。 (未经同行评审认证)是作者/资助者。 本文是美国政府作品。 它不受 17 USC 版权的约束。 此预印本的版权持有者此版本于 2022 年 3 月 19 日发布。;https://doi.org/10.1101/2022.03.18.484947 doi:bioRxiv 预印本
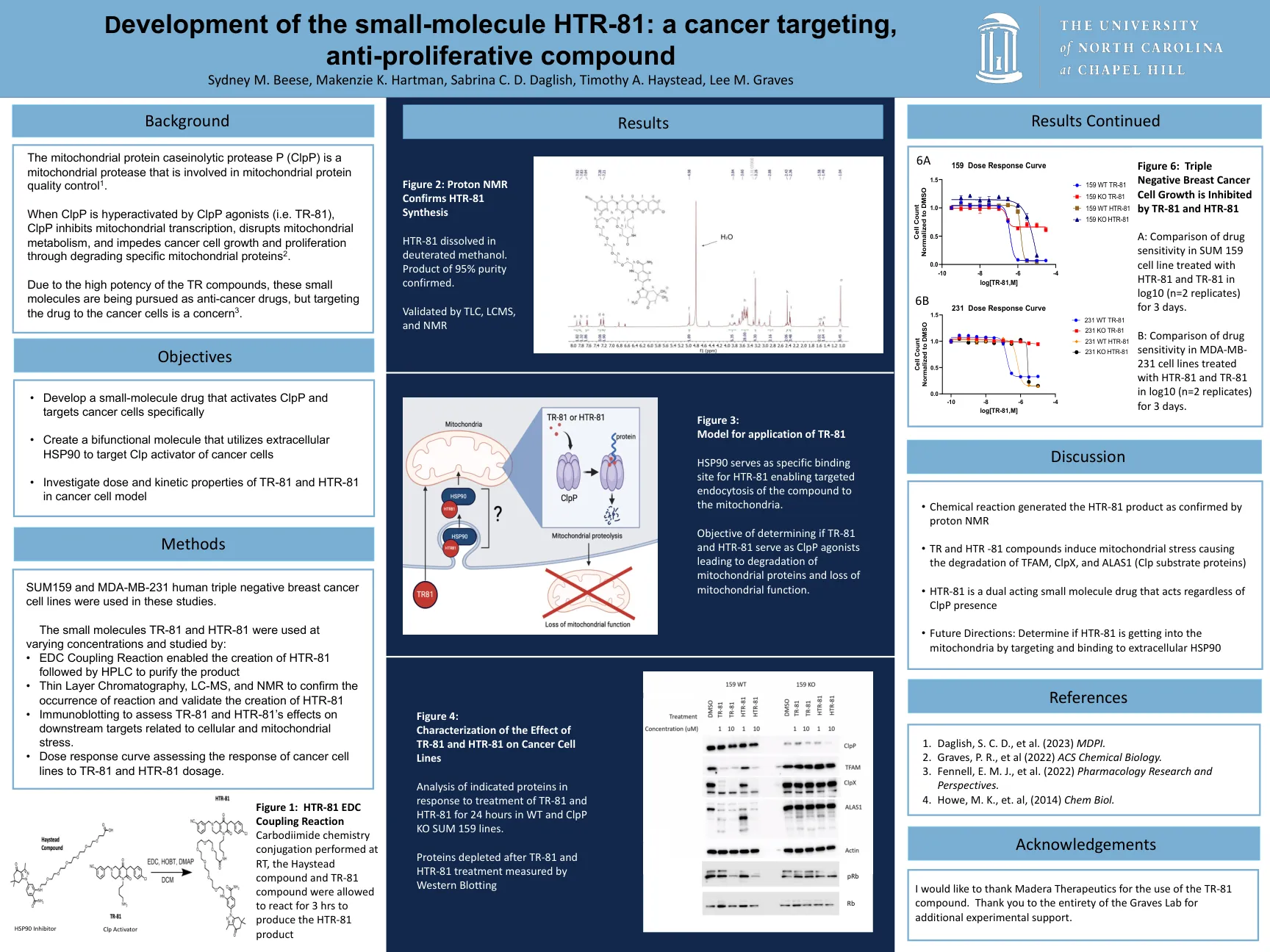
小分子 HTR-81 的开发:一种抗癌药物……
当 ClpP 被 ClpP 激动剂(即 TR-81)过度激活时,ClpP 会抑制线粒体转录,破坏线粒体代谢,并通过降解特定的线粒体蛋白 2 来阻碍癌细胞的生长和增殖。


叶绿体基因组完整分析:东方山羊豆的结构、系统发育关系及山羊豆科植物的进化推断
摘要:东方山羊豆是豆科植物,具有重要的生态和经济价值,因其抗逆性强、蛋白质含量高而被广泛栽培。然而,东方山羊豆的基因组信息尚未见报道,限制了其进化分析。由于基因组较小,叶绿体相对容易获得基因组序列以进行系统发育研究和分子标记开发。本文对东方山羊豆叶绿体基因组进行了测序和注释。结果表明,东方山羊豆叶绿体基因组长度为125,280 bp,GC含量为34.11%。共鉴定出107个基因,包括74个蛋白质编码基因,29个tRNA和4个rRNA。东方山羊豆叶绿体基因组中丢失了一个反向重复(IR)区。此外,与其近缘种G. officinalis的叶绿体基因组相比,有5个基因( rpl22 、 ycf2 、 rps16 、 trnE-UUC 和 pbf1 )丢失。共检测到84个长重复序列和68个简单序列重复序列,可作为G. orientalis及其近缘种遗传研究的潜在标记。我们发现,在G. officinalis与其他3个Galegeae物种( Calophaca sinica 、 Caragana jubata 、 Caragana korshinskii )的两两比较中,petL 、 rpl20 和 ycf4 3个基因的Ka/Ks值大于1,表明这3个基因受到了正向选择。 15个Galegeae物种的比较基因组分析表明,大多数保守的非编码序列区域和两个基因区域(ycf1和clpP)分化程度较高,可作为DNA条形码用于快速准确的物种鉴定。基于ycf1和clpP基因构建的系统发育树证实了Galegeae物种间的进化关系。此外,在所分析的15个Galegeae物种中,Galega orientalis在ycf1基因中有一个独特的30 bp内含子,而Tibetan liangshanensis在clpP基因中缺少两个内含子,这与现有只有IR缺失支(IRLC)中的甘草属物种缺少两个内含子的结论相反。总之,首次确定并注释了G. orientalis的完整叶绿体基因组,这可以为Galegeae属内尚未解决的进化关系提供见解。
在Wendelstein 7-X
引言维持蛋白质稳态(蛋白质稳定)对于正常的细胞功能至关重要,并且蛋白质失调的蛋白抑制剂与许多类型的癌症有关(1-3)。蛋白质症是由未折叠的蛋白质反应(UPR)调节的,该蛋白质反应(UPR)在内质网和线粒体中被激活以减轻各种细胞应激(4-6)。线粒体UPR(UPR MT)促进细胞适应普遍的线粒体应激。线粒体特异性伴侣和蛋白酶的UPR MT作用,以主导线粒体质量控制(3,7-10)。线粒体伴侣 - 一种活性对于在线粒体中正确折叠和展开的蛋白质的折叠至关重要。两个伴侣系统,热休克蛋白60(HSP60)和线粒体HSP70(MTHSP70),促进线粒体基质(11-16)中的蛋白质折叠功能。相关,MTHSP70与HSP10(HSP60的辅助因子)合作,以促进成熟HSP60复合物的组装(17)。在哺乳动物细胞中已经鉴定出超过26个线粒体蛋白酶,其中LON肽酶1(LONP1)和酪蛋白溶解蛋白酶P(CLPP)发挥了突出作用(6)。这些蛋白酶降解
CLP Power Hong Kong Limited碳中立性...
可持续性,并鼓励我们的业务伙伴和供应商在供应链管理方面采用最佳实践,并确保良好的企业公民身份。我们负责任的采购政策声明(REPP)总结了CLPP在4个关键领域(或“要素”)中对供应商实践的要求和期望:法律合规,尊重人(包括劳动/工作条件和安全性),道德与商业行为,道德与商业行为以及环境管理。- CLP Power根据其对业务运营,环境,社会和治理(ESG)风险和总合同承诺的重要性来定义“关键项目”。对于这些关键项目,供应商通过各种工具(包括自称问卷,提案评估和现场访问)评估其可持续性实践。-CLP Power已为购买类别开发了绿色规格,以支持日常操作。我们致力于提高对可持续消费的认识。如果绿色采购要素是重要的考虑因素,CLP通常会对整个采购过程进行整体审查,以确保可持续的要求

靶向 Clp 蛋白酶的 BacPROTAC:一种有前途的抗分枝杆菌药物研发策略
两千年来,结核病 (TB) 夺走的生命比世界上任何其他传染病都要多。2021 年,世界卫生组织 (WHO) 估计有 1060 万人被诊断出患有结核病,导致 140 万 HIV 阴性患者死亡。耐多药结核病 (MDR-TB)(定义为至少对利福平 (RIF) 和异烟肼 (INH) 具有耐药性)和广泛耐药结核病 (XDR-TB) 的出现是未来几年要克服的主要挑战。我们最近对该领域的投资和研究工作进行了广泛的分析,总体目标是到 2030 年实现消除结核病的既定里程碑。在过去几年中,在将多种有前景的化合物推进到临床开发阶段方面取得了显着进展,每种化合物都具有不同的作用机制。但值得注意的是,已经出现了对目前某些抗结核药物产生耐药性的分枝杆菌菌株。创新的蛋白水解靶标嵌合体 (PROTACs) 蛋白质降解方法的探索已成为发现新型抗菌药物的可行途径。虽然泛素系统是真核细胞独有的,但某些细菌使用类似的降解系统,该系统依赖于 ClpC:ClpP (ClpCP) 蛋白酶对磷酸化精氨酸残基 (pArg) 的识别,从而导致蛋白质降解。在这篇评论文章中,我们描述和分析了利用细菌蛋白水解机制 (BacPROTACs) 的 PROTACs 设计新型抗结核药物的进展。范围声明。由于耐药菌株的出现,开发用于治疗结核病的新型药物被认为迫切需要。在此背景下,引进能够减轻疾病并实现世界卫生组织所概述的目标的新技术势在必行。在创新策略中,降解对杆菌生存至关重要的蛋白质有望产生新药物,特别是那些对治疗潜伏(非复制性)结核分枝杆菌有效的药物。从这个角度来看,我们介绍了结核分枝杆菌治疗领域取得的进展和遇到的障碍

转基因生物:牛津疫苗组(20...
B2 与申请有关的转基因生物的一般描述 GMO 的描述 GMO 是野生型副伤寒沙门氏菌 (S. Paratyphi) A 9150 菌株的同源突变体。遗传改造的目的是构建一种带有 guaBA 操纵子和 clpX 基因缺失的改良 S. Paratyphi A 9150 菌株,以产生生长缺陷的减毒 S. Paratyphi A 菌株 (CVD 1902)。GMO (CVD 1902) 将用于研究其作为减毒活口服疫苗在受控人类感染模型中预防肠热病的价值。CVD 1902 副伤寒沙门氏菌 A 血清型活口服疫苗是由野生型亲本菌株 S. Paratyphi A 9150 构建的。使用改良的 Lambda Red 介导的定点诱变程序进行缺失。删除了两个基因序列:guaBA 染色体操纵子(编码远端从头鸟嘌呤核苷酸生物合成途径中使用的两种酶)和 clpX 基因(编码分子伴侣 ATPase,与 clpP 编码的丝氨酸蛋白酶一起发挥作用,形成参与各种代谢过程的复合物)。clpX 缺失突变的表型后果之一是鞭毛的过度表达,这也可能增强 GMO 菌株的免疫原性,因此有助于其作为减毒活疫苗的适用性。从研究中获得的信息将用于指导疫苗设计和开发,从而可能影响公共卫生干预策略。应用描述在这项临床研究中,我们建议调查 GMO CVD 1902 作为减毒活疫苗在 S. Paratyphi A 人类攻击模型中预防副伤寒感染的有效性。使用野生型 S 的 S. Paratyphi A 感染的人感染模型。副伤寒甲型(NVGH308 株)已在牛津疫苗组(英国牛津大学)建立。牛津疫苗组(英国牛津大学)一直在进行