XiaoMi-AI文件搜索系统
World File Search SystemFret

发育期 Pb 暴露通过改变 Hipsc 衍生皮质神经元的细胞内 Ca2+ 稳态增加 AD 风险
在脆弱的发育时期接触铅 (Pb) 等环境化学物质会导致晚年健康出现不良后果。人类队列研究表明,发育期 Pb 暴露与晚年阿尔茨海默病 (AD) 发病之间存在关联,动物研究的结果进一步证实了这一观点。然而,发育期 Pb 暴露与 AD 风险增加之间的分子通路仍然难以捉摸。在这项工作中,我们使用人类 iPSC 衍生的皮质神经元作为模型系统来研究 Pb 暴露对人类皮质神经元中 AD 样发病机制的影响。我们将来自人类 iPSC 的神经祖细胞暴露于 0、15 和 50 ppb Pb 中 48 小时,去除含 Pb 的培养基,并进一步将它们分化为皮质神经元。免疫荧光、蛋白质印迹、RNA 测序、ELISA 和 FRET 报告细胞系用于确定分化皮质神经元中 AD 样发病机制的变化。将神经祖细胞暴露于低剂量 Pb,模拟发育暴露,可导致神经突形态改变。分化神经元表现出钙稳态、突触可塑性和表观遗传景观的改变,以及 AD 样发病机制标志物升高,包括磷酸化 tau、tau 聚集体和 A β 42/40。总之,我们的研究结果为发育性 Pb 暴露引起的 Ca 失调提供了证据基础,这是一种合理的分子机制,可解释发育性 Pb 暴露人群中 AD 风险的增加。

磷酸盐感应器细胞器调节磷酸盐和组织稳态
无机磷酸盐(P I)是生命的必需分子之一。然而,对动物组织中的细胞内P I代谢和信号传导知之甚少。在观察到慢性P I饥饿会导致果蝇的消化性上皮中引起过度增殖,我们确定P I饥饿会触发P I Transporter PXO的下调。与P I饥饿一致,PXO缺乏引起中肠过增高。有趣的是,免疫染色和超微结构分析表明,PXO特异性标记了非典型的多层细胞器(PXO主体)。此外,通过使用Förster共振能量转移(FRET)P I传感器2进行P i成像,我们发现PXO限制了胞质P I水平。PXO身体需要PXO进行生物发生,并在P I饥饿后发生降解。PXO体的蛋白质组学和脂质组表征揭示了其独特的特征,作为细胞内P I储备。因此,P I饥饿会触发PXO下调和PXO体降解,作为增加胞质P I的补偿机制。最后,我们将激酶的连接器与AP-1(CKA)(CKA)(CKA)和JNK信号3的一个组件(CKA)确定为PXO敲低或P I饥饿诱导的高增殖的介体。总的来说,我们的研究将PXO体作为胞质P I水平的关键调节剂,并鉴定出P i依赖性的PXO – CKA – JNK信号传导控制组织稳态。

靶向肌球蛋白结合蛋白C
抽象心脏MYBP-C(CMYBP-C)与肌动蛋白 - 肌球蛋白相互作用,以微调心脏肌肉收缩。CMYBP-C的磷酸化降低了CMYBP-C与肌动蛋白或肌球蛋白的结合,通常会减少心力衰竭(HF)患者,并且在HF模型系统中受到心脏保护。因此,CMYBP-C是模仿磷酸化和/或扰动其与肌动蛋白或肌球蛋白相互作用的HF药物的潜在靶标。我们用荧光素-5-甲酰亚胺(FMAL)和CMYBP-C(CC0-C2)的C0-C2片段标记了肌动蛋白(CC0-C2)。我们在FDA批准的药物库上进行了两个互补的高通量筛选(HTS),以发现专门结合CMYBP-C的小分子,并使用荧光寿命(FLT)检测来影响其与肌动蛋白或肌球蛋白的相互作用。我们首先激发了FMAL并检测到其FLT,以测量荧光共振能量转移(FRET)的变化,从FMAL(供体)到TMR(受体),表明蛋白质复合物中的结合和/或结构变化。使用相同的样品,然后使用更长的波长激光直接激发TMR,以检测化合物对TMR环境敏感FLT的影响,以识别直接结合CC0-C2的化合物。次要测定,对主要HTS分析的选定调节剂进行,这些化合物的特异性是磷酸化对未磷酸化的CC0-C2的特异性,而CC0-C2对C1-C2与C1-C2相对于快速骨骼肌的C1-C2(FSKC1-C2)。确定的化合物的子集调制了心脏和/或骨骼肌原纤维中ATPase活性。简介这些测定法确定了发现CMYBP-C-肌动蛋白/肌球蛋白相互作用的小分子调节剂的可行性,其最终目的是开发HF疗法。

用双标记的自我快速,全细胞DNA-Paint成像...
1。Schnitzbauer,J。等。用DNA-Paint的超分辨率显微镜。nat。原始。12.6,1198-1228(2017)。 2。 Reinhardt,S。C.等。 Ångström-分辨率荧光显微镜。 自然617.7962,711-716(2023)。 3。 Jungmann,R。等。 用DNA-Paint和Exchange-Paint多路复用3D细胞超分辨率成像。 nat。 方法11.3,313-318(2014)。 4。 Auer,A。 使用基于FRET的探针快速,无背景的DNA绘制成像。 纳米lett。 17.10,6428-6434(2017)。 5。 Chung,K。K.等。 荧光DNA-Paint,可更快,低背景超分辨率成像。 nat。 方法19.5,554-559(2022)。 6。 Knemeyer,J。P。等。 基于染料二聚化的自淬灭DNA探针,用于鉴定分枝杆菌。 int。 J. Environ。 肛门。 化学。 85,625-637(2005)。 7。 Kessler,L。F.等。 自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。 angew。 化学。 int。 ed。 Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。12.6,1198-1228(2017)。2。Reinhardt,S。C.等。Ångström-分辨率荧光显微镜。自然617.7962,711-716(2023)。3。Jungmann,R。等。 用DNA-Paint和Exchange-Paint多路复用3D细胞超分辨率成像。 nat。 方法11.3,313-318(2014)。 4。 Auer,A。 使用基于FRET的探针快速,无背景的DNA绘制成像。 纳米lett。 17.10,6428-6434(2017)。 5。 Chung,K。K.等。 荧光DNA-Paint,可更快,低背景超分辨率成像。 nat。 方法19.5,554-559(2022)。 6。 Knemeyer,J。P。等。 基于染料二聚化的自淬灭DNA探针,用于鉴定分枝杆菌。 int。 J. Environ。 肛门。 化学。 85,625-637(2005)。 7。 Kessler,L。F.等。 自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。 angew。 化学。 int。 ed。 Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。Jungmann,R。等。用DNA-Paint和Exchange-Paint多路复用3D细胞超分辨率成像。nat。方法11.3,313-318(2014)。4。Auer,A。使用基于FRET的探针快速,无背景的DNA绘制成像。纳米lett。17.10,6428-6434(2017)。5。Chung,K。K.等。 荧光DNA-Paint,可更快,低背景超分辨率成像。 nat。 方法19.5,554-559(2022)。 6。 Knemeyer,J。P。等。 基于染料二聚化的自淬灭DNA探针,用于鉴定分枝杆菌。 int。 J. Environ。 肛门。 化学。 85,625-637(2005)。 7。 Kessler,L。F.等。 自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。 angew。 化学。 int。 ed。 Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。Chung,K。K.等。荧光DNA-Paint,可更快,低背景超分辨率成像。nat。方法19.5,554-559(2022)。6。Knemeyer,J。P。等。基于染料二聚化的自淬灭DNA探针,用于鉴定分枝杆菌。int。J. Environ。肛门。化学。85,625-637(2005)。 7。 Kessler,L。F.等。 自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。 angew。 化学。 int。 ed。 Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。85,625-637(2005)。7。Kessler,L。F.等。 自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。 angew。 化学。 int。 ed。 Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。Kessler,L。F.等。自淬灭的荧光团二聚体,用于DNA涂料和st型显微镜。angew。化学。int。ed。Engl。 E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。Engl。E202307538(2023)。 8。 Bollmann,S。等。 有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。 物理。 化学。 化学。 物理。 13.28,12874-12882(2011)。 9。 nat。 10。 nat。E202307538(2023)。8。Bollmann,S。等。有机荧光团的二聚体形成在变性条件下报告了生物分子动力学。物理。化学。化学。物理。13.28,12874-12882(2011)。9。nat。10。nat。Mortensen,K。I.,Churchman,L。S.,Spudich,J.A.和Flyvbjerg,H。单分子跟踪和超分辨率显微镜的优化定位分析。方法,7.5,377-381(2010)。Helmerich,D。A.等。 照片处理指纹分析绕过10 nm的分辨率障碍。 方法19.8,986-994(2022)。 11。 Huisken,J。等。 通过选择性平面照明显微镜在实时胚胎深处的光学切片。 科学305.5686,1007-1009(2004)。Helmerich,D。A.等。照片处理指纹分析绕过10 nm的分辨率障碍。方法19.8,986-994(2022)。11。Huisken,J。等。通过选择性平面照明显微镜在实时胚胎深处的光学切片。科学305.5686,1007-1009(2004)。

SNCF 集团 2021 12 财务报告
JEAN-PIERRE FARANDOU SNCF 集团董事长兼首席执行官 « 2021 年较 2020 年出现了大幅反弹,尤其是在下半年,客运量在夏季和年底再次反弹。我们的 2021 年业绩使我们有望履行对法国政府(我们的唯一股东)的财务承诺,即到 2022 年实现正自由现金流。我们在 2021 年下半年做到了这一点,这要归功于我们高度适应的团队、对业务的积极管理以及持续的结构性成本削减,从而使自由现金流恢复盈利。法国政府的这种严格管理和支持使我们能够推进对铁路系统整体绩效至关重要的项目:调整运输计划、改善服务基础(安全性、可靠性和乘客信息)以及开展重建和升级铁路网络和基础设施的工作。这些结果还表明,我们的多元化战略是正确的选择。GEODIS 报告了创纪录的收入,这得益于全球货运和电子商务的强劲增长。KEOLIS 的增长巩固了 SNCF 集团在可持续交通综合解决方案方面的全球领导地位。Fret SNCF 实现收支平衡——这对欧洲铁路货运的未来来说是一个非常令人鼓舞的迹象。2021 年,我们看到了开放客运市场竞争的第一个具体效果。SNCF RÉSEAU 和 SNCF GARES & CONNEXIONS 现在正本着公平公正的精神与新参与者合作,SNCF VOYAGEURS 的产品和服务也更具竞争力。SNCF 集团业务遍及法国全境和全球 120 个国家,我们制定了雄心勃勃的企业社会责任路线图,以实现整体业绩的持久增长。我们必须继续推进业务转型和创新,使 SNCF 集团成为可持续交通领域的全球领导者。我要向 SNCF 集团在法国和全球的所有团队致以最诚挚的谢意。我对他们在整个疫情期间的出色参与表示敬意 »

鉴定针对黄素病毒和冠状病毒的蛋白酶抑制剂
载体传播的黄病毒和人畜共患病的冠状病毒是重要的人类病原体,对全球公共卫生构成了严重威胁。黄病毒,在那里它们被节肢动物载体传播,每年引起数百万个感染。虽然大多数感染是轻度或无症状的,但登革热和黄热病病毒(如黄热病病毒)可能会引起潜在的致命性出血热和休克综合征。神经性黄病毒,例如西尼罗河,日本脑炎和tick虫脑炎(TBEV)会导致具有长期症状的脑膜脑炎。冠状病毒,尤其是SARS(2003)和MERS(2012)(2012年)(2012年)的人畜共患病毒,如2000年代初期一直在定期出现。最新的例子是SARS-COV-2,它在中国城市武汉引起了一系列感染后,遍布全世界,目前造成超过690万人死亡。尽管在SARS-COV-2的情况下,疫苗对于预防感染或严重疾病和住院是至关重要的,但抗病毒药是一种非常有价值的工具,可用于治疗和预防当前和未来的黄病毒和冠状病毒感染。在本文中提出的工作中,我们使用了硅和体外技术的组合来识别和测试病毒蛋白酶潜在抑制剂的活性。在我们的第一项研究(论文1)中,我们意外地鉴定出具有对ZIKV NS2B-NS3蛋白酶的体外活性的HIV蛋白酶抑制剂。Covid-19爆发后,我们将注意力转移到SARS-COV-2上。通过虚拟筛选已知蛋白酶抑制剂的库来鉴定抑制剂,该抑制剂通过分子动力学模拟评估,并最终使用基于FRET的酶促测定法对重组ZIKV蛋白酶进行了测试。还使用了分子对接和分子动力学模拟的相同组合来正确预测已知的泛氟韦蛋白酶抑制剂对TBEV蛋白酶的活性(论文2)。结果,我们是第一个报告基于肽的化合物,具有对TBEV的体外活性。我们首先测试了广谱抗病毒一氧化氮(NO)的抑制作用,并发现无释放的化合物快照对基于细胞的测定中的SARS-COV-2复制具有剂量依赖性抑制作用(论文3)。我们推测SNAP可以通过对SARS-COV-2主要蛋白酶的催化Cys145的反硝化来抑制SARS-COV-2蛋白酶,并发现SNAP对体外酶试验中的重组SARS-COV-2 MPRO蛋白酶具有剂量依赖性抑制作用。在我们的最后一项研究(论文4)中,我们通过对含有42亿种化合物的DNA编码的化学文库的亲和力筛选来确定一类新的有效SARS-COV-2蛋白酶抑制剂。所鉴定的化合物抑制了IC50低至25 nm的重组SARS-COV-2蛋白酶,并且在感染的CALU-3和CACO-2细胞系中低微摩尔范围内的剂量依赖性抗病毒作用。

朝着电动汽车电池的互动管理
多个PEG链的水合体积。TX100是一种表面活性剂,具有乙氧基甲氧基辛基的基本骨架,带有一个亲水头和一个疏水性尾巴的长矛状结构。使用荧光光谱法检查了表面活性剂与模型抗原之间的相互作用,据说这比UV-VIS光谱,5和NMR光谱谱比敏感性高1000倍,该光谱具有与UV-VIS光谱的敏感性相当的敏感性。牛血清白蛋白(BSA)长期以来一直详细研究了溶液中的抗原性和抗原性,被选为模型抗原。6,7我们还专注于环糊精(CD)作为抗原疏水核心的通用模型,因为长期以来一直将CD作为酶的底物结合位点的模型研究,从1954年的Einschlussverbindunger(包含化合物)出版。8有一些使用CD衍生物作为氧化酶和酯酶模型的例子。9,10最近,据报道CD衍生物是脂肪酶的模型,这些脂肪酶可以选择性地水解疏水腔中的溶血磷脂。11因此,CD在历史上被认为是酶的底物结合位点的模型,这是外部疏水物质界面的典型示例,并探索辅助表面活性剂在其上的作用如何被认为是理想的实验系统,可以普遍地模拟蛋白质的疏水核心核心核心。在这项研究中,在环脱糖蛋白中选择了羟丙基-B-环糊精(HP-B -CD),该研究具有明确定义的疏水性和疏水性表面,并最大程度地显示了疏水性荧光探针的荧光(见下文)。使用特定的蛋白质,例如BSA,卵蛋白(OVA)和核糖核酸酶(RNase)作为抗原模型,不允许我们摆脱其独特的特性,12并利用CD作为抗原核心核心的模型,可以为这个问题提供解决方案。通过评估疏水性荧光探针与模型抗原疏水性核心的吸附和结合,评估了各种非离子表面活性剂与模型抗原BSA和HP -B -CD模型抗原之间的相互作用。The hydrophobic core environment of BSA and HP- b -CD was evaluated by the fluorescence of 8-anilinonaphthalene-1- sulfonic acid (ANS), a hydrophobic fluorescent probe whose fluorescence is enhanced in hydrophobic environments or adsorbed in the lipid bilayer of liposomes, in the hydrophobic core of proteins, 13–17 or in the表面活性剂的胶束。18因此,ANS用于评估这些大分子和小分子提供的疏水环境。然而,一定浓度后,ANS和其他荧光分子的荧光强度开始降低。这称为浓度猝灭,由于内部滤波器效应,它被广泛称为淬火。19其他可能的淬火机制包括forster共振能量转移(FRET)和DEXTER机制,20,21是由荧光分子彼此接近造成的。无论机制如何,荧光分子数量增加引起的淬火是评估中培养基和大分子提供的疏水环境的障碍。为了解决这个问题,我们在本研究中利用了抑制剂模型。

开放博士学位学生 /博士后职位:HIV-1大会和成熟电话:2024年5月28日,传染病系,海德尔伯>病毒学< / div>
开放博士学位学生 /博士后职位:HIV-1组装和成熟会致电:2024年5月28日,海德堡大学传染病系,病毒学系为PhD学生提供职位,并在所有职业生涯水平上为逆转录病毒复制中的组装,成熟和入场后的活动提供职位。研究将在DFG资助的合作研究中心1129(https://www.sfb1129.de/)的Hans-GeorgKräusslich和BarbaraMüller组中进行。我们的小组研究了HIV-1组装的多个方面,成熟以及进入以及逆转录病毒复制的早期阶段。HIV-1组装发生在专门的膜微区域生产者细胞的质膜上。颗粒最初以不成熟的非感染形式释放。与颗粒释放,病毒蛋白酶对主要结构多蛋白的蛋白水解裂解导致病毒形态的急剧改变。然而,蛋白酶激活和调节的机理以及成熟过程仍然很少理解。成熟的,锥形的衣壳最近被公认为是HIV-1复制后进入后事件的关键编排(Müller等,2022)。我们正在研究颗粒形成的结构和动态方面,特别关注蛋白酶激活和随后的变化(Schimer等,2015; Hanne等,2016; Qu等人。2021)。复杂的组装,成熟和进入后事件通过传统批量方法逃脱分析。因此,我们使用高级显微镜以高时间或空间分辨率可视化单个粒子水平的事件。与Briggs组合作获得的数据(Qu et al。,2021)令人惊讶地揭示了不仅帽子蛋白晶格,而且还表明,矩阵(MA)晶格在蛋白水解成熟后都会进行结构重排,这表明MA在现实事件中目前不知名的作用。这涉及活细胞成像,FRET/FLIM,Sted和MinFlux纳米镜以及相关的光和电子显微镜和冷冻电子显微镜,后者与其他组合作。这些研究将与病毒学和细胞生物学方法以及计算模拟相辅相成,以研究蛋白酶激活和成熟的过程及其生物学后果。我们在互动和国际科学环境中提供了具有生物医学相关性的退出和高度跨学科的研究主题,包括在国际竞争性的水平上与国家和国际合作伙伴的合作。这些项目是SFB1129的一部分,涉及研究中心内不同学科的群体的密切互动。实验室位于综合传染病研究中心(www.ciid-heidelberg.de),该研究还设有独特的最新IDIP成像平台(www.idip-heidelberg.org)。成功的博士生候选人将入学HBIGS国际研究生院(http://www.hbigs.uni-heidelberg.de/),以从该计划的出色科学培训中受益。PostDocs将有资格获得Heidelberg University Heitracks计划(https://www.uni-heidelberg.de/en/research/support-for-early-career-career-cresearch-careearcher-careercearchers/carifate-support)的结构化职业支持。申请人应具有相关学科的硕士学位或博士学位(分子和细胞生物学,生物化学,生物物理学或分子医学)。他们应该有兴趣使用各种方法解决基本的病毒学问题,并使用
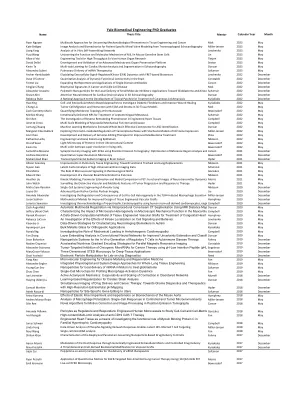
耶鲁大学生物医学工程博士毕业生
Ryan Nguyen 用于揭示组织工程和癌症中的机械生物学现象的多尺度方法 Mak 2023 年 5 月 Kate Bridges 经食道超声心动图患者特定二尖瓣建模的图像分析和生物力学 Miller-Jensen 2023 年 5 月 Liang Yang 体外自组装网络的分析 Levchenko 2023 年 5 月 Yuqi Wang 揭示小鼠生殖系干细胞中 MILI 的功能和分子机制 Lin 2023 年 5 月 Alborz Feizi 用于高通量离体人体器官研究的工程工具 Tietjen 2023 年 5 月 David Dellal 先进机电器官保存平台的开发和验证 Sestan 2023 年 5 月 Kevin Ta 超声心动图心脏运动分析和分割的多任务学习 Duncan 2023 年 5 月 Alexandra Suberi mRNA 治疗的肺部递送 Saltzman 2023 年 5 月 Archer Hamidzadeh 使用基于 FRET 的生物传感器阐明细胞外信号调节激酶 (ERK) 动力学 Levchenko 2022 年 12 月 Dave O'Connor 脑内动态功能连接的定量分析 Constable 2022 年 12 月 Feimei Liu 扩展单域抗体库和应用 Carson 2022 年 12 月 Xingjian Zhang 癌症和镰状细胞病的生物物理特征 Mak 2022 年 12 月 Alexander Josowitz 用于局部递送小分子抑制剂的聚合物纳米粒子:胶质母细胞瘤和气道的应用 Saltzman 2022 年 12 月 Shawn Ahn 注意力神经网络在 3D 超声心动图心脏应变分析中的应用 Duncan 2022 年 12 月 Rebecca Byler 治疗皮肤利什曼病的局部贴剂开发的合理方法 Kyriakides 2022 年 12 月 Hao Xing 基于细胞和细胞外基质的方法研究糖尿病成纤维细胞并改善伤口愈合 Kyriakides 2022 年 5 月 Chang Liu 3D 组织模型中肿瘤细胞的迁移以及与 ECM 和基质的相互作用 Mak 2022 年 5 月 Zach Connerty-Marin 在纳米尺度上量化膜拓扑结构 Bewersdorf 2022 年 5 月 MinSoo Khang 鞘内递送 NP 用于治疗软脑膜转移 Saltzman 2022 年 5 月 Shi Shen 逆转录病毒的研究工程心脏组织中的重塑现象 Campbell 2022 年 5 月 Jenette Creso 心肌机械功能和疾病的多尺度建模 Campbell 2022 年 5 月 Juntang Zhuang 机器学习方法估计全脑有效连接组以识别自闭症 Duncan 2022 年 5 月 Margaret Elise Bullock 使用 HIV 基因表达随机模型探索染色质介导的转录噪声调控 Miller-Jensen 2022 年 5 月 Ann Chen 开发和提供基因组编辑疗法以改善胶质母细胞瘤治疗 Zhou 2022 年 5 月 Katherine Leiby 工程功能性远端肺上皮 Niklason 2022 年 5 月 Ons M'Saad 蛋白质在其超微结构背景下的光学显微镜检查 Bewersdorf 2022 年 5 月 Kevin Hu 活细胞中的多色各向同性超分辨率 Bewersdorf 2022 年 5 月 Samantha Rossano Synaptic使用正电子发射断层扫描的 SV2A 密度成像:参考区域分析的优化和 Carson 2021 年 12 月 Andrew Barentine 定量超分辨率显微镜 Bewersdorf 2021 年 12 月 Muhammad Khan 脑癌跨室钠成像 Hyder 2021 年 12 月 Allison Greaney 肺组织工程的改进:迈向功能性气管和肺置换 Niklason 2021 年 5 月 Siyuan Gao 高维脑成像数据的潜在因子分析 Scheinost 2021 年 5 月 Rita Matta 微血管信号在神经源性微环境的作用 Gonzalez 2021 年 5 月 Edward Han 血管生物人工内分泌胰腺的开发 Niklason 2021 年 5 月 Heather Liu PET 中的动力学建模、参数估计和模型比较:神经递质动力学的功能图像 Morris 2021 年 5 月 John Walsh 监测肿瘤进展和治疗反应的独特血管和代谢特征 Hyder 2021 年 5 月 Micha Sam Raredon 肺泡肺的单细胞系统工程 Niklason 2020 年 12 月 Luyao Shi 高级定量心脏核成像 Liu 2020 年 12 月 Amanda Alexander 研究 TLR4 诱导的巨噬细胞分泌中细胞间异质性的调节和后果 Miller-Jensen 2020 年 12 月 Jason Szafron 用于改进组织工程血管移植物设计的数学模型 Humphrey 2020 年 12 月 Lorenzo Sewanan 使用人类干细胞衍生的心肌细胞、enginCampbell 2020 年 12 月 Zach Augenfeld 自动使用 MRI 距离图通过术中锥形束 CT 分割进行多模态配准 Duncan 2020 年 5 月 Jeffery (Alex) Clark 表征微尺度异质性对心肌宏观机械功能的影响 u Campbell 2020 年 5 月 Ramak Khosravi 用于治疗先天性心脏病的组织工程血管移植物的数据驱动计算模型 D Humphrey 2020 年 5 月 Rebecca LaCroix 激酶定位对细胞信号传导和行为影响的研究 Levchenko 2020 年 5 月 Xiaoxiao Li 用于表征自闭症神经影像生物标志物的数据驱动策略 Duncan 2020 年 5 月 Ayomiposi Loye 用于骨科应用的块状金属玻璃 Kyriakides 2020 年 5 月 Ronald Ng 研究机械负荷在致心律失常性心肌病中的作用 Campbell 2020 年 5 月 Fan Zhang Layer卷积神经网络中的嵌入分析可改善不确定性估计和分类 Duncan 2020 年 5 月 Sean Bickerton 纳米粒子系统用于在体内生成调节性 T 细胞用于自身免疫性疾病治疗 Fahmy 2019 年 12 月 Nadine Dispenza 加速非线性梯度编码策略用于并行磁共振成像 Constable 2019 年 12 月 Alexander Svoronos 使用 pH 低插入肽 (pHL) 进行肿瘤靶向抑制致癌微小 RNA 用于癌症治疗 Engelman 2019 年 12 月 MaryGrace Velasco 用于深层组织应用的三维 STED 显微镜 Bewersdorf 2019 年 12 月 Shari Yosinski 用于片上实验室诊断的电子粒子操作 Reed 2019 年 12 月 Yang Xiao 微血管工程用于疾病建模和再生医学 Fan 2019 年 5 月 Alexander Engler 综合生理与系统设计全肺组织工程方法 Niklason 2019 年 5 月 Young-Eun Seo 用于局部递送 miRNA 抑制剂治疗胶质母细胞瘤的纳米粒子 Saltzman 2019 年 5 月 Zhuo Chen 用于分析巨噬细胞活化动力学的单细胞微芯片 Fan 2019 年 5 月 Ian Linsmeier 活性肌动球蛋白力学:无序网络中收缩的协同性和缩放性 Murrell 2018 年 12 月 Haiying (Allen) Lu 基于学习的心脏应变分析正则化 Duncan 2018 年 12 月