XiaoMi-AI文件搜索系统
World File Search SystemGATA1

2024指南
可遇到另外的细胞减少症(中性粒细胞减少症比血小板减少症的频率更高),婴儿的短暂性血小板病。2除了患有GATA1突变的病例。3强烈暗示DBA综合征;但是不够具体,无法进行诊断。仅在专业实验室中进行4个研究测试;在模棱两可或非信息遗传学的情况下有用。 5通常在成年人中呈现。 6这些IBMF通常表现出多型细胞减少症,并且经常出现其他影响多器官系统的疾病特异性异常。 这种区别特征可以帮助将这些条件与DBA综合征区分开,该疾病最初以孤立的红细胞发育不全表现出来。 缩写:BM;骨髓; EADA,红细胞腺苷脱氨酶; HBF,胎儿血红蛋白; SLE,全身性红斑狼疮; prca,纯红细胞性植物; CLL,慢性淋巴细胞性白血病; LGL,大颗粒状淋巴细胞性白血病; CT,计算机断层扫描; MRI,磁共振成像; IBMF,继承的骨髓衰竭综合征; FA,Fanconi贫血; SDS,Shwachman钻石综合征; DC,Dyskeratosis Congenita。仅在专业实验室中进行4个研究测试;在模棱两可或非信息遗传学的情况下有用。5通常在成年人中呈现。6这些IBMF通常表现出多型细胞减少症,并且经常出现其他影响多器官系统的疾病特异性异常。这种区别特征可以帮助将这些条件与DBA综合征区分开,该疾病最初以孤立的红细胞发育不全表现出来。缩写:BM;骨髓; EADA,红细胞腺苷脱氨酶; HBF,胎儿血红蛋白; SLE,全身性红斑狼疮; prca,纯红细胞性植物; CLL,慢性淋巴细胞性白血病; LGL,大颗粒状淋巴细胞性白血病; CT,计算机断层扫描; MRI,磁共振成像; IBMF,继承的骨髓衰竭综合征; FA,Fanconi贫血; SDS,Shwachman钻石综合征; DC,Dyskeratosis Congenita。

双向表观遗传编辑揭示基因调控的层次结构
CRISPR 扰动是研究基因组功能效应的宝贵工具。然而,现有方法在研究非编码元件和遗传相互作用方面的效用有限。在这里,我们开发了一个双向表观遗传编辑系统 (CRISPRai),其中正交激活 (CRISPRa) 和抑制 (CRISPRi) 扰动同时应用于同一细胞的多个基因座。我们开发了双 gRNA 捕获单细胞 Perturb-seq 来研究两种造血谱系转录因子 SPI1 和 GATA1 之间已建立的相互作用,并发现了共同调节基因的新型上下文特定调控模式。将 CRISPRai 扩展到非编码元件,我们解决了多个增强子如何相互作用以调节 T 细胞中共同靶基因白细胞介素-2 的表达。我们发现增强子功能主要是附加的并能够对基因表达进行微调,但在基因表达控制强度方面,增强子之间存在明显的层次结构。启动子在控制基因表达方面比大多数增强子占主导地位;然而,一小部分增强子表现出强大的功能效应或守门人功能,尽管启动子被激活,但仍可以关闭基因。将这些功能数据与组蛋白 ChIP-seq 和 TF 基序富集相结合,表明存在多种增强子介导的基因调控模式。我们的方法 CRISPRai 用于双向表观遗传编辑,提供了一种识别新遗传相互作用的方法,这些相互作用在没有双向扰动的情况下进行研究时可能会被忽视,并且可以应用于基因和非编码元件。

一种新型遗传病、幼年型粒单核细胞白血病和骨髓增生性肿瘤中的 SH2B3 变异
造血是人类生命周期中不断发展的高度动态过程。胎儿期和围产期是特殊的生理变化和进化时期。在这些发育阶段,体质性遗传条件可能导致维持和分化干细胞和祖细胞的有效调节剂失衡,从而导致新生儿和幼儿出现特定的血液学表型。一个突出的例子是唐氏综合征,其 21 三体性介导的胎儿造血紊乱。大约 10% 的唐氏综合征新生儿出现暂时性异常骨髓造血,其特征是外周血原始细胞增多和转录因子 GATA1 的特异性体细胞突变。1 虽然 10% 至 20% 的病例在出生后 4 年内转变为全面性白血病,但大多数病例无需治疗即可痊愈。另一种常见的发育障碍是努南综合征,该综合征由 RAS/丝裂原活化蛋白激酶 (MAPK) 通路的种系致病变异引起,可在出生后头几个月表现为暂时性骨髓增生性疾病 (MPD)。大多数努南综合征患者携带 PTPN11 种系突变。2 尽管唐氏综合征中暂时性异常髓系生成的突变情况至少已得到部分阐明,但努南综合征相关 MPD 的机制仍然很大程度上不清楚。Perez-Garcia 等人 3 和 Blombery 等人 4 报道了另一种暂时性 MPD,发生在 SH2B3 基因双等位基因种系突变的患者出生后不久。 SH2B3 编码淋巴细胞衔接子 LNK(也称为 SH2B3),是 SH2B 衔接子蛋白家族的成员,该家族还包括 APS(SH2B2)和 SH2B(SH2B1)(图 1)。SH2B 蛋白具有共同的结构,即 N 端二聚化结构域、中央 pleckstrin 同源性 (PH) 结构域和 C 端 Src 同源性

一种重写γ-球蛋白启动子并将胎儿血红蛋白重新激活的主要编辑策略
1。Frangoul,H。等。exagamglogene自动赛,用于严重的镰状细胞疾病。n Engl J Med 390,1649–1662(2024)。2。忘记,B。G。胎儿血红蛋白的遗传持久性的分子基础。ann。N. Y. Acad。 SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。N. Y. Acad。SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。SCI。850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。850,38–44(1998)。3。Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。KLF1在英国HPFH中驱动胎儿血红蛋白的表达。血液130,803–807(2017)。4。Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。NAT COMUM 6,7085(2015)。5。Martyn,G。E.等。近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。血液133,852–856(2019)。6。Martyn,G。E.等。自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。nat Genet 50,498–503(2018)。7。Frati,G。等。CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。8。Anzalone,A。V。等。搜索和重新固定基因组编辑,无需双链断裂或供体DNA。自然576,149–157(2019)。9。Coleman,M。B.等。am。J. Hematol。42,186–190(1993)。 10。 Chen,P。J.等。42,186–190(1993)。10。Chen,P。J.等。Chen,P。J.等。g伽玛A伽马(β+)胎儿血红蛋白的遗传持久性:g伽玛-158 c-> t在顺式中与-175 t-> c c gamma-lobin基因的突变会导致G Gama-- gamma基因的增加导致G Gama-Globobin的增加。通过操纵细胞决定因素的编辑结果来增强质量编辑系统。Cell 184,5635-5652.E29(2021)。 11。 Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Cell 184,5635-5652.E29(2021)。11。Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Ravi,N。S.等。通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。Elife 11,E65421(2022)。12。Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Kim,H。K.等。预测人类细胞中主要编辑指南RNA的效率。nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。13。Nelson,J。W.等。设计的Pegrnas提高了主要的编辑效率。NAT生物技术40,402–410(2022)。14。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。核酸Res 50,1187–1197(2022)。15。Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Lee,J。等。prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。nat Commun 14,1786(2023)。16。Antoniou,P。等。基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。nat Commun 13,6618(2022)。17。Pavani,G。等。通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。血液Adv 5,1137–1153(2021)。18。Everette,K。A.等。在体内造血干细胞的体内质量编辑促进小鼠植入后镰状细胞疾病表型。nat Biomed Eng 7,616–628(2023)。19。Peterka,M。等。利用DSB修复以促进有效的同源性依赖性和 - 独立的质量编辑。nat Commun 13,1240(2022)。20。Magnani,A。等。对镰状细胞疾病的同种异体移植后混合嵌合体患者进行了广泛的多系数分析:对基因治疗的造血和植入阈值的见解。Haematologica 105,1240–1247(2020)。21。Sun,Y。等。 在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Sun,Y。等。在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。在小鼠中耐用基因校正的肺部干细胞的体内编辑。科学384,1196–1202(2024)。22。Doman,J。L.等。噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。单元格186,3983-4002.E26(2023)。23。Wimberger,S。等。同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。nat Commun 14,4761(2023)。24。Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Yan,J。等。用内源性的小RNA结合蛋白改善原始编辑。自然628,639–647(2024)。25。Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。26。核酸res。Brinkman,E。K.,Chen,T.,Amendola,M。&Van Steensel,B。通过序列痕量分解对基因组编辑的易于定量评估。42,E168(2014)。 27。 Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和42,E168(2014)。27。Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和Brusson,M。等。新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。mol the核酸32,229–246(2023)。28。Gaudelli,N。M.等。腺嘌呤基础编辑者的定向演变,活动增加和
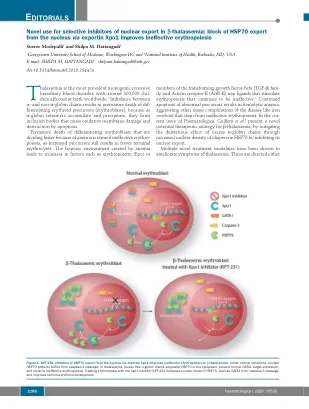
社论
改善未结合的A -Globin和非A -Globin链之间的平衡或纠正无效的红细胞。修饰的TFG-β家族受体拮抗剂,如Sotatercemp(ACE-011)和Luspatercept(ACE-536)阻止配体与ACTR-II受体结合,并随后激活SMAD4信号通路,4改善Ery-Throid Throid Cell和红细胞的产生。通过CRISPR Therapeatics成功的基因疗法实现了未结合A -Globin与非A -Globin链的异常比例,并得到了波士顿顶点药物的支持。称为CTX001的体细胞疗法使用了编辑的患者自己的造血干细胞(HSC)来刺激胎儿血红蛋白的产生。5通过XPO1抑制HSP70的细胞内局部局部局部局部可能会合并这两个治疗目标。几条证据表明,红细胞使用分子伴侣在红细胞发育过程中对不稳定的过量A -Glo- bin链分割,6-8,因此,靶向这种伴侣的靶向时,当过量的globobin tetramer会累积时,靶向这种伴侣可能在β -tha -thaplamasemia中有用。许多组指出,分子伴侣HSP70在红细胞9-11中积聚至高水平,对于简化胚芽成熟很重要。11正常的人红细胞成熟需要在成熟后期的caspase-3瞬时激活,以防止过度的红细胞生产。激活的胱天蛋白酶可以切割GATA-1,从而导致成熟停滞和/或凋亡。12 Ribeil等。 14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。12 Ribeil等。14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。表明EPO会导致HSP70转移到核中,结合GATA-1并保护其免受caspase-3裂解。相反,在EPO剥夺期间,HSP70被排除在细胞核中,而GATA-1被cas-pase-3裂解,导致凋亡死亡。13因此,HSP70的细胞内位置的改变似乎在红细胞生存力中起关键作用(图1)。在β-丘脑贫血中观察到的无效性红细胞生成的特征是在多染色体阶段加速了红细胞分化,成熟停滞和凋亡。在人β-thal虫蛋白粒细胞的成熟过程中,HSP70直接通过过量的游离A-格珠蛋白链直接在细胞质中螯合(图1)。核定靶向的HSP70突变体或caspase-3-无分解的GATA-1突变体恢复了β-thal核阿无血成红细胞的终末成熟。14在Haematologica,Guillem等。3跟进这种机制,以表明导出蛋白1(XPO1)调节在正常条件下HSP70在红细胞中的HSP70的核质质位置。Guillem等。证实,用XPO1抑制剂KPT-251治疗红细胞增加了HSP70的核水平,从caspase-3裂解中救出了GATA1,并改善了末端红细胞原理(图1)。尽管使用核出口的选择性抑制剂(SINE)用于治疗淋巴瘤和多骨髓瘤