XiaoMi-AI文件搜索系统
World File Search SystemGATA2

产生两个杂合的 GATA2 CRISPR/Cas9-...
GATA2 缺陷属于世界卫生组织 (WHO) 新近确定的一组易患髓系恶性肿瘤的遗传综合征(Smith et al., 2004)。具有种系杂合 GATA2 突变的个体表现出非常复杂和多系统的表现型,包括血细胞减少导致的 MDS、免疫缺陷(涉及 B、NK、单核细胞、CD4 +、DC 细胞谱系)、耳聋和淋巴水肿(Hahn et al., 2011)。根据文献报道,至少 75% 的 GATA2 突变携带者在估计的中位年龄 20 岁时患上 MDS/AML(Wlodarski et al., 2016)。如今,化疗和同种异体造血干细胞 (HSC) 移植仍然是唯一具有良好反应的治疗方法。由于缺乏可靠的疾病模型系统,我们无法从机制上理解 GATA2 单倍体不足如何影响造血发育。种系 GATA2 突变要么是截短的功能丧失 (LOF) 突变,要么是 ZF2 的错义突变,要么是破坏内含子 4 增强子位点的突变 ( Wlodarski et al., 2016 )。这些突变被认为会导致 GATA2 功能降低/丧失,特别是消除 ZF2 的 DNA 结合功能 ( Chong et al., 2018 )。迄今为止,只有少数种系 GATA2 突变进行了功能研究。因此,使用精确的基因编辑策略,我们生成了携带两种最

表观基因组分析显示,GATA2缺乏症中异常的DNA甲基化特征
GATA2表达是一种复杂的多系统疾病,患有骨髓增生综合征(MDS)和急性髓样白血病(AML)具有高风险,并且寿命几乎完全完整。1,2 GATA2载体表现出高度可变的表达性,一些人在早期发作MD中发育,而其他人则在整个生命中仍然无症状。尽管不存在预后生物标志物,但合作遗传和表观遗传驱动因素可能会塑造疾病的进程。3尽管在一组白血病驱动基因(即Stag2,setBP1,ASXL1和ETV6)中鉴定出复发的体细胞突变的进展,但了解GATA22携带者中与白血病相关的molecu Lar机制存在重大差距。4此外,DNA甲基化改变有助于在成人PA和AML的成年PA含量中发生白血病克隆和异常高甲基化的启动和扩展。5,6迄今为止,尚未进行GATA2患者的基因组宽DNA甲基分析。在这项研究中,招募了7家西班牙医院的20个临床注释的GATA2载体(表1;在线SUP培养图S1)。诊断时中位年龄为36(范围6-75)。主要初始表现为MDS(n = 12,55%),其次是免疫效率(n = 3,15%)和AML(n = 2,10%)。在细胞遗传学上,三个患者中的三体术在两个患者中进行了复杂的核型,而有11例患者的核型正常(在线补充图S1A-F)。基于DNA的可用性,因此在17 GATA2载体的总外周血(Pb)或骨髓(BM)中进行了乳酸突变。因此,在71%(12/17)测试的患者中鉴定了髓样恶性基因中的乳酸突变(在线补充图S1F)。该分析证实了GATA2缺乏效率中获得的体细胞突变的异质性,stag2,asxl1和setBP1作为经常影响的基因。,我们获得了通过使用矿体内甲基化史诗850 K平台(Illumina)进行完全遗传表征的患者的全球DNA甲基化pro纤维。我们为八个BM(P1,P3,P5,P6,P10,P11,P11,P12,P13)和八个PB样品(P1,P1,P4,P7,P7,P8,P9,P9,P13,P13,P16和P17)的LED DNA进行了比较,并与12(5 pb和7 BM)的队列进行了比较(5 pb和7 bm)老年人(Hefleped Donor)(hded donor)(高维数据可视化表明,GATA2患者的射击性紧密聚集在一起,并从HD隔离。属于同一家族的无症状载体P1,P1,P16和P17(分别为6、40和75岁)被包含在HD组上(图1A)。此外,我们将HD与无症状

crispr/cas9基因编辑在血压茎中clolls clollal clollal竞争,用于GATA2 Defenscy Remero-Moya*,Oskar
CRISPR/Cas9 Gene Editing in Hematopoietic Stem Cells to Model Clonal Competition in vivo and in vitro for GATA2 Deficiency Damia Romero-Moya*, Oskar Marin-Bejar, Maximiliano Distefano, Joan Pera, Jessica Gonzales, Arnau Iglesias, Marcin Wlordaski, Anna Bigas, Alessandra Giorgetti.再生医学计划,D'EnvessiveAcióbiomèdicade Bellvitge(IDIBELL),医院te llobregat,西班牙GATA2缺乏症已被确定为儿童骨髓质发育综合综合症(MDS)和儿童急性骨髓性白血病的常见遗传性原因。受影响家庭中的外观和表现力通常是可变的,这表明需要合作因素触发疾病。MDS驱动基因中的体细胞突变(SETBP1,ASXL1)已被鉴定出GATA2-MDS。触发GATA2载体中白血病进展的分子机制仍然未知。特别是这些问题仍然没有解决:1)GATA2种系突变本身是否足以触发MDS/AML 2)SETBP1和ASXL1突变是否诱导恶性转化。由于缺乏忠实的人类疾病模型,解决这些问题非常困难。在这里,我研究了单独携带GATA2突变的体外/体内工程脐带血CD34+细胞,或与NSG小鼠中的SETBP1/ASXL1突变结合使用,以评估植入能力和克隆进化。具体来说,CRISPR/CAS9/RAAV6用于在CD34+细胞(单个)中引入R398W突变,或与SETBP1和ASXL1突变(多路复用)一起引入R398W突变。在所有条件下,主要和次要移植都显示出相似的多核构成。有趣的是,遗传研究表明,在多重条件下主要扩展的克隆携带了SETBP1+ASXL1突变,而仅丢失了GATA2突变的克隆。为了研究转录组水平的突变的影响,SCRNASEQ正在进行中。体外数据证实,携带GATA2 R398W突变的细胞具有损害的克隆能力和增殖,从而概括了MDS患者表型。总而言之,我们通过针对CD34+细胞的CRISPR/CAS9开发了人类的克隆竞争模型。我们的发现强烈表明,GATA2 R398W突变不足以增加细胞适应性,这表明遗传,表观遗传学,利基和压力因子的合作是触发疾病所必需的。
uyr_gata2_final_042224.pdf
GATA2缺乏会导致免疫系统问题,使您更容易感染。GATA2缺乏的其他常见特征包括一种称为肺肺泡蛋白质病,过度肿胀(淋巴水肿)的肺部疾病,以及血细胞数量减少。血细胞的降低会导致一种称为骨髓增生综合征(MDS)的疾病,该疾病可以发展为急性髓样白血病(AML)。但是,GATA2缺乏症可以在身体的许多部位出现多种症状。

使用临床外显子组测序
先天性免疫力(IEI)包括多种异质遗传疾病,其中免疫系统中的缺陷导致对感染和其他并发症的敏感性增加。准确,及时诊断IEI对于治疗计划和预后至关重要。在这项研究中,评估了临床外显子组测序(CE)诊断IEI的临床实用性。对于37例与IEI相关的症状,体征或实验室异常的韩国患者,CES涵盖了4,894个基因,包括与IEI相关的基因。审查了他们的临床诊断,临床特征,感染家族病史和实验室结果以及检测到的变体。使用CES,在37例患者中有15例(40.5%)对IEI进行了遗传诊断。 从IEI相关的基因,BTK,UNC13D,STAT3,IL2RG,IL2RG,IL10RA,NRAS,SH2D1A,GATA2,TET2,TET2,PRF1和UBA1中检测到了17种致病变异,其中四种变体先前是一致的。 其中,从GATA2,TET2和UBA1中鉴定出体细胞病变变体。 此外,我们通过CES偶然诊断出了两名患者,该患者是为了诊断未识别的IEI患者的其他疾病而进行的。 综上所述,这些结果证明了CES诊断为IEI的实用性,这有助于准确的诊断和适当的治疗。使用CES,在37例患者中有15例(40.5%)对IEI进行了遗传诊断。从IEI相关的基因,BTK,UNC13D,STAT3,IL2RG,IL2RG,IL10RA,NRAS,SH2D1A,GATA2,TET2,TET2,PRF1和UBA1中检测到了17种致病变异,其中四种变体先前是一致的。其中,从GATA2,TET2和UBA1中鉴定出体细胞病变变体。此外,我们通过CES偶然诊断出了两名患者,该患者是为了诊断未识别的IEI患者的其他疾病而进行的。综上所述,这些结果证明了CES诊断为IEI的实用性,这有助于准确的诊断和适当的治疗。

DYRK1A 和 GATA1 在 21 三体巨核细胞生成中的协同作用
简介:患有唐氏综合症 (DS) 或 21 三体综合症 (T21) 的儿童罹患暂时性异常髓系造血 (TAM) 和唐氏综合症急性巨核细胞白血病 (ML-DS) 的风险较高 (1, 2)。TAM 是一种新生儿前白血病,由胎儿时期 T21 与 GATA1s 的独特遗传相互作用引起,GATA1s 是关键造血转录因子 GATA 结合蛋白 1 (GATA1) 的 N 端截短异构体。TAM 和 ML-DS 母细胞均以 GATA1 体细胞突变为特征,从而产生 GATA1s (3, 4),但 ML-DS 母细胞还会获得“第三次打击”突变,通常是在表观遗传调节因子或黏连蛋白复合物成员中 (5, 6)。值得注意的是,在缺乏 T21 的个体中,生殖细胞 GATA1s 突变会导致先天性贫血、血小板减少和/或中性粒细胞减少,但与白血病无关 (7, 8),这证实了 GATA1s 和 T21 共同促进白血病的必要性。细胞周期在造血发育过程中受到精确控制。GATA1 已被证实能抑制细胞周期进程和增殖,并通过阻止转录激活因子 E2Fs 与其下游靶标结合来促进造血细胞的终末分化 (9–11)。Rb/E2F 通路对细胞周期调控至关重要,通常受 GATA1 抑制;然而,由于 GATA1 N 端对这种相互作用至关重要,GATA1s 无法抑制激活因子 E2Fs (9–11)。 GATA1 还抑制 GATA2(GATA 结合蛋白 2),GATA2 是一种造血转录因子,对造血干细胞 (HSC) 和巨核细胞扩增至关重要,在 ML-DS 中经常过表达 (12)。由于没有 N 端结构域,GATA1s 无法正确下调 GATA2,导致 HSC 和巨核细胞过度增殖 (13, 14)。

社论:利用计算机方法发现抗癌药物的最新趋势
大量的药物研发工作集中于调节与癌症相关的分子通路,特别是涉及功能或表达改变的蛋白质的通路,这也为个性化医疗奠定了基础。在这种情况下,计算方法极大地支持了药物发现过程( Rosales-Hernandez 等人,2012 年;Lavecchia 和 Di Giovanni,2013 年;Cerchia 和 Lavecchia,2023 年;Romanelli 等人,2024 年),在某些情况下代表了发现新型小分子疗法背后的驱动力。本研究主题收集了宝贵的贡献,展示了抗癌药物发现领域的成功案例,并强调了与变革性计算机模拟方法的协同作用。Marção 等人的研究文章报道了使用一类逻辑回归机器学习算法(OCLR)来估计人类癌细胞的干性,还提供了各种犬和乳腺癌细胞的干性指数。干细胞特性与癌症的发生和发展以及恶性肿瘤和治疗耐药性有关。随后,作者研究了小分子 (+)-JQ1 对溴结构域和末端外 (BET) 蛋白家族抑制的影响,以及随之而来的犬癌细胞干细胞特性抑制。通过分析公开数据,他们注意到人类三阴性乳腺癌细胞系 MDA-MB- 231 中也有类似的结果,从而指出通过 BET 抑制进行表观遗传调控是解决干细胞特性的一个有趣策略。这项研究为机器学习在理解化合物对癌症干细胞特性的影响方面的潜力提供了宝贵的见解,这对于开发靶向疗法至关重要。在 Menendez-Gonzalez 等人的文章中,作者描述了通过基于配体的虚拟筛选发现一种新型 GATA2 抑制剂,为急性髓细胞白血病 (AML) 治疗提供了一个有希望的途径。该研究通过基于形状的筛选和分子对接来识别潜在的 GATA2 抑制剂,随后进行体外和体内实验来评估对 AML 细胞的抑制作用。已识别的 GATA2 抑制剂化合物 11 带有取代的哌嗪骨架,通过诱导细胞凋亡和抑制细胞增殖表现出强大的抗 AML 活性。
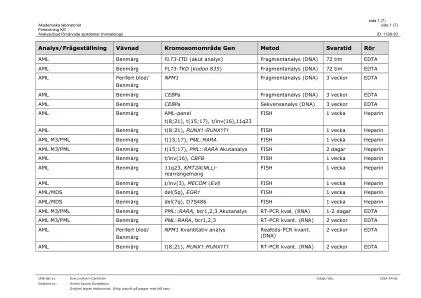
分析提供获得的疾病(血液学)
leukemiutringbenmärg/blod mylood面板*(ABL1,ANKRD26,ASXL1,ATRX,BCOR,BCOR,BCORL1,BRAF,CALR,CALR,CBL,CBL,CBL,CBL,CDKN2A,CDKN2A,CEBPA,CEBPA,CEBPA,CSF3R,CSF3R,CSF3R,CUX1,DDX41,DDX41,DNMT3A fbxw7, FLT3, GATA1, GATA2, GNAS, HRAS, Idh1, Idh2, Ikzf1, jak2, jak3, kdm6a, kit, kraas, kmt2a, mpl, myd88, NF1, Notch1 (INKLUSIVE 3´UTR), NPM1, NRAS, PDGFRA, PHF6, PPM1D, Pten, Ptpn11, Rad21, Runx1, Samd9, SAMDL9, Setbp1, SF3B1, SMC1A, SMC3, SRSF2, Stag2, Stat3, Stat5B, Tet2, TP53, U2AF1, WT1, ZRSR2, BTK, plcg2, terc) Div>

AD 2 - LCRA - 1 - 1 英国军用 AIP 阿克罗蒂里
1. 注意。10/28 号跑道的混凝土末端和距离 10 号跑道入口 2000 英尺以内的跑道区域在潮湿时容易打滑,特别是如果发现雨后有积水。 2. 注意。鸟击风险高,特别是在春秋两季的迁徙季节。 3. 电缆 插图:a. 10 号跑道 - 373 米/1223 英尺。b. 28 号跑道 - 374 米/1227 英尺。正常操作 - 两条电缆均已拆除。快速喷气机操作 - App 电缆已拆除,超限电缆已升起。 4. 电路 a. 方向。10 号跑道 RHC:28 号跑道 LHC。b. 高度。(i)正常 - 1100 QNH。(ii)重度 - 1600 QNH。 (iii) FJ 低空转弯 - 600 QNH。5. TKOF 后保持 Rwy Tr 直到飞过海面。6. 附加频率:Talkdown 240·05、GATA1 240·1、GATA2 355·0、APS Ops 369·45、SAR 252·8。

EVI1控制KDM6B介导的组蛋白去甲基化为...
等离子体,单核细胞,中性粒细胞或血小板的增殖增加(1、3、4)。大约30%的被诊断为MD的患者最终患有急性髓样白血病(AML)(5)。eVI1首先被鉴定为具有逆转录病毒诱导的髓样恶质的小鼠中生态病毒整合的常见位点(6)。人类EVI1(MECOM)基因位于Chro-Mosome 3Q26上,EVI1的多种同工型在MECOM基因座(7)中编码。3q26染色体的重排,导致EVI1的上调,经常发生在包括MDS,AML和慢性髓样白血病(CML)在内的髓样恶性疾病中(8-10)。MDS,AML和CML具有INV(3)/T(3; 3)重排通常具有相似的病理特征,预后不良(8、11、12)。It was reported that chromosome rear- rangements cause overexpression of EVI1 due to relocation of enhancers, including GATA binding protein 2 (GATA2) enhancer in inv(3)/t(3;3) (q21q26) (13, 14) and MYC super-enhancer in t(3;8) (q26;q24) close to the EVI1 gene (15).EVI1过表达可能发生在没有3染色体重排的MDS患者中。EVI1上调