XiaoMi-AI文件搜索系统
World File Search SystemHaematologica

Bernhard R. Gentner
Scopus作者ID:6507111312。Orcid ID: http://orcid.org/0000-0001-6024-4718 Selected publications (* shared first/last authorship) Caserta C*, Nucera S*, […], Gentner B. miR-126 identifies a quiescent and chemo-resistant human B-ALL cell subset that correlates with minimal residual disease.白血病。2023年10月; 37(10):1994-2005。 doi:10.1038/s41375-023-02009-5。PMID:37640845 Naldini MM,[…],Gentner B.急性髓样白血病中化学疗法反应的纵向单细胞分析。nat Commun。2023 3月8日; 14(1):1285。Gentner B等人,用于hur综合征的造血茎和祖细胞基因疗法。n Engl J Med。2021 11月18日; 385(21):1929-1940。 doi:10.1056/nejmoa2106596。PMID:34788506。影响因子:91.25。引用:2。现场加权引用影响:2.27。Capo V,[…],Gentner B&Villa A.扩展了循环造血干/祖细胞,作为治疗TCIRG1骨质骨异常的新细胞来源。血液学。2021 1月1日; 106(1):74-86。 doi:10.3324/haematol.2019.238261。PMID:31949009。影响因素:9.94。引用:7。field-w。引文影响:2.90。Zonari E,[…],Gentner B.有效的离体工程和高度纯化的人造血茎和祖细胞群的基因疗法的扩展。干细胞报告。2017年4月11日; 8(4):977-990。 doi:10.1016/j.stemcr.2017.02.010。PMID:28330619。影响因素:7.77。引用:87。field-w。引文影响:5.19。Chiriaco M等人,Gentner B*,Aiuti A*。X连锁慢性

pirtobrutinib:带有柔软侧面的“蛮力”
In this issue of Haematologica , Shah and colleagues report toxicity and outcomes of a subgroup of 127/597 patients with low-grade B-cell lymphomas, previously intolerant to at least one Bruton tyrosine kinase inhibitor (BTKi) and without progressive disease (PD), who were treated with pirtobrutinib in the phase I/II BRUIN study.1此亚组分析提供了临床相关的数据,因为尽管发生不良事件(AE),但尽管临床疗效持续进行临床功效,但多达40%的接受共价BTKI的患者仍会停止治疗。这样的AE包括心房颤动,出血,感染,腹泻,皮疹和关节痛,其中2个是由于BTKI与EGFR,SRC和TEC(例如EGFR,SRC和TEC)的结合。3折叠曲替尼是一种非共价BTKI,BTK的选择性> 100倍,与363/370(98%)的其他激酶进行了测试,从而限制了远不含量的毒性。4作为原理证明,折叠式曲丁替尼与先前发表的复发 /耐火性慢性淋巴细胞白细胞(CLL) /小细胞淋巴细胞淋巴瘤(sll)和麦芽瘤的患者(<5%)的停药率相关。5,6在Shah及其同事的研究中,有94%的患者以前在其疾病病程中的某个时候接受了依鲁替尼。 雌性丁比的感染的一半是相关的,因为大多数研究都是在19009年大流行期间进行的。 这突出了预防性的重要性5,6在Shah及其同事的研究中,有94%的患者以前在其疾病病程中的某个时候接受了依鲁替尼。雌性丁比的感染的一半是相关的,因为大多数研究都是在19009年大流行期间进行的。这突出了预防性的重要性1导致伊布鲁蒂NIB中断的最常见不良事件是心脏疾病(32%),感染(10%),中性粒细胞减少症(9%),皮疹(9%),肢数 /肌痛(8%),出血 /出血(7%),胃部疾病(6%)(6%)(5%)(5%),(5%),(5%),(6%),(6%),(5%),(5%),(5%),(5%),(5%),(5%)(5%),(5%)(5%)(5%)与CLL/SLL中ibrutinib的其他研究一致。7,8总共有64%的治疗生气AE导致先前的BTKI中断并未与折叠式折叠抗体发生,但除了感染,中性粒细胞减少症和胃肠道疾病外,这些复发率> 50%,尽管这些因素> 50%,但这些疾病> 50%,但由于不适合于2例患者,因此这些疾病率低或散发出了2个患者的症状。
社论
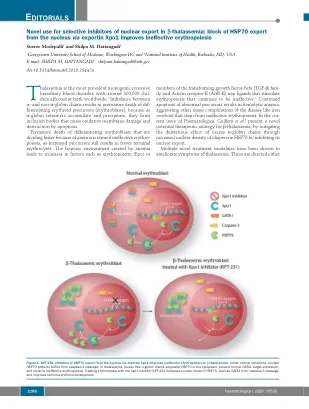
改善未结合的A -Globin和非A -Globin链之间的平衡或纠正无效的红细胞。修饰的TFG-β家族受体拮抗剂,如Sotatercemp(ACE-011)和Luspatercept(ACE-536)阻止配体与ACTR-II受体结合,并随后激活SMAD4信号通路,4改善Ery-Throid Throid Cell和红细胞的产生。通过CRISPR Therapeatics成功的基因疗法实现了未结合A -Globin与非A -Globin链的异常比例,并得到了波士顿顶点药物的支持。称为CTX001的体细胞疗法使用了编辑的患者自己的造血干细胞(HSC)来刺激胎儿血红蛋白的产生。5通过XPO1抑制HSP70的细胞内局部局部局部局部可能会合并这两个治疗目标。几条证据表明,红细胞使用分子伴侣在红细胞发育过程中对不稳定的过量A -Glo- bin链分割,6-8,因此,靶向这种伴侣的靶向时,当过量的globobin tetramer会累积时,靶向这种伴侣可能在β -tha -thaplamasemia中有用。许多组指出,分子伴侣HSP70在红细胞9-11中积聚至高水平,对于简化胚芽成熟很重要。11正常的人红细胞成熟需要在成熟后期的caspase-3瞬时激活,以防止过度的红细胞生产。激活的胱天蛋白酶可以切割GATA-1,从而导致成熟停滞和/或凋亡。12 Ribeil等。 14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。12 Ribeil等。14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。表明EPO会导致HSP70转移到核中,结合GATA-1并保护其免受caspase-3裂解。相反,在EPO剥夺期间,HSP70被排除在细胞核中,而GATA-1被cas-pase-3裂解,导致凋亡死亡。13因此,HSP70的细胞内位置的改变似乎在红细胞生存力中起关键作用(图1)。在β-丘脑贫血中观察到的无效性红细胞生成的特征是在多染色体阶段加速了红细胞分化,成熟停滞和凋亡。在人β-thal虫蛋白粒细胞的成熟过程中,HSP70直接通过过量的游离A-格珠蛋白链直接在细胞质中螯合(图1)。核定靶向的HSP70突变体或caspase-3-无分解的GATA-1突变体恢复了β-thal核阿无血成红细胞的终末成熟。14在Haematologica,Guillem等。3跟进这种机制,以表明导出蛋白1(XPO1)调节在正常条件下HSP70在红细胞中的HSP70的核质质位置。Guillem等。证实,用XPO1抑制剂KPT-251治疗红细胞增加了HSP70的核水平,从caspase-3裂解中救出了GATA1,并改善了末端红细胞原理(图1)。尽管使用核出口的选择性抑制剂(SINE)用于治疗淋巴瘤和多骨髓瘤

isatuximab,pomalidomide和地塞米松作为打捞
恩里卡·安东尼亚·马蒂诺(Enrica Antonia Martino),丹尼尔·德鲁达斯(Daniele Derudas),埃琳娜·罗西(Elena Rossi),索菲亚·特里兹(Sofia Terlizzi),乔瓦尼(Giovanni) O,Roberta Della Pepa,Giuseppe Bertuglia,Emiliano Barbieri,Conchitale,Conconce。 Claudio de Magistris,Lorenzo de Paoli,Velia Bongarzoni,Anna Maria Cafro,Anna Mele,Pietro Benvenuti,Claudio Cerchione,Cirino Botta,Elisabetta Antoniooli Bianco,Massimiliano Aragio Fabbo,Gloria Margio Casaluci, Angela Rago, Raffaele Fontana, Francesca Farina, Ernesto Vigna, Antonella Bruzzese, Katia Mancuso, Davide Nappi, Sonia Morè, Elena Rivati, Catello Califano, Angela Amendola, Daniela Roccotelli, Alessandra Lombardo, Annalisa Citro, Giuseppina Bartcello, Renato Zambello, Alessandro Maggi, Santo Neri, Michele Monachesi, Alessandro Gozzetti, Vittorio Montefusco, Vittorio Montefusco. Marino Bruori,Emilia Cotzia,Giuseppe Pietrantuono,Angela Maria Quinto,Valeria Amico,Nicola di Renzo,Marta Coscia,Monica Galli,Valerio de Stefano,Maria Teresa Petrucci,Antonino neri neri ando, :2024年9月15日。接受:2024年11月11日。 iSatuximab,pomalidomide和地塞米松作为患者多发性骨髓瘤的挽救疗法:意大利,多中心,回顾性临床经验,在对照临床试验之外,有270例病例。 血液学。 2024年11月28日。doi:10.3324/haematol.2024.286658 [Epub在印刷前]出版商的免责声明。iSatuximab,pomalidomide和地塞米松作为患者多发性骨髓瘤的挽救疗法:意大利,多中心,回顾性临床经验,在对照临床试验之外,有270例病例。血液学。2024年11月28日。doi:10.3324/haematol.2024.286658 [Epub在印刷前]出版商的免责声明。引用:Enrica Antonia Martino,Daniele Derudas,Elena Rossi,Sofia Terlizzi,Giovanni Reddicto,Paola Stefanoni,Jacopo Micozzi,Silvia Mangiacavali Aro,Roberta Della Pepa,Giuseppe Bertuglia,Emiliano Barbieri,Concetta Concetta Concetta Corticello,Claudio de Magistris,Lorenzo de Paoli,Velia Bongarzoni,Anna Maria Cafro,Anna Mele,Anna Mele,Pietro Benvenuti,Claudio Cerchione,Cirino Botta,Elisabetta botta,Elisabetta Rosario Bianco,Massimiliano Arangio Fabbo,Gloria Margiotta,Gloria Margiotta。 Casaluci, Angela Rago, Raffaele Fontana, Francesca Farina, Ernesto Vigna, Antonella Bruzzese, Katia Mancuso, Davide Nappi, Sonia Morè, Elena Rivati, Catello Califano, Angela Amendola, Daniela Roccotelli, Alessandra Lombardo, Annalisa Citro, Giuseppina Bird, Renato Zambello, Alessandro Maggi, Santo Neri, Michele Monachetti, Vittorio, Montefusco,Marino Bruori,Emilia Cotzia,Giuseppe Pietrantuono,Angela Maria Quinto,Valeria Amico,Nicola di Renzo,Marta Coscia,Monica Coscia,Monica Galli,Valerio de Stefano,Maria teresa petrucci,anterino neriboto,forto dii dii dii dii dii Massimo Gentile。e-发布在印刷前对于快速传播科学至关重要。因此,所有适用于该期刊的法律免责声明也与该生产过程有关。在印刷之前发行电子版本后,手稿将进行技术和英语编辑,排版,证明校正和呈现以供作者的最终批准;然后,手稿的最终版本将出现在日记的常规期刊中。

一种重写γ-球蛋白启动子并将胎儿血红蛋白重新激活的主要编辑策略
1。Frangoul,H。等。exagamglogene自动赛,用于严重的镰状细胞疾病。n Engl J Med 390,1649–1662(2024)。2。忘记,B。G。胎儿血红蛋白的遗传持久性的分子基础。ann。N. Y. Acad。 SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。N. Y. Acad。SCI。 850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。SCI。850,38–44(1998)。 3。 Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。850,38–44(1998)。3。Wienert,B。等。 KLF1在英国HPFH中驱动胎儿血红蛋白的表达。 血液130,803–807(2017)。 4。 Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。KLF1在英国HPFH中驱动胎儿血红蛋白的表达。血液130,803–807(2017)。4。Wienert,B。等。 编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。 NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。Wienert,B。等。编辑基因组,以引入与胎儿球蛋白增加有关的有益天然发生的突变。NAT COMUM 6,7085(2015)。 5。 Martyn,G。E.等。 近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。 血液133,852–856(2019)。 6。 Martyn,G。E.等。 自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。 nat Genet 50,498–503(2018)。 7。 Frati,G。等。 CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。 mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。 8。 Anzalone,A。V。等。 搜索和重新固定基因组编辑,无需双链断裂或供体DNA。 自然576,149–157(2019)。 9。 Coleman,M。B.等。 am。 J. Hematol。 42,186–190(1993)。 10。 Chen,P。J.等。NAT COMUM 6,7085(2015)。5。Martyn,G。E.等。近端启动子中的自然调节突变通过创建从头GATA1部位来提高胎儿球蛋白表达。血液133,852–856(2019)。6。Martyn,G。E.等。自然调节突变通过破坏BCL11A或ZBTB7A结合来提升胎儿球蛋白基因。nat Genet 50,498–503(2018)。7。Frati,G。等。CRISPR-CAS9治疗镰状细胞病的安全性和功效研究突出了特异性疾病的反应。mol ther s1525-0016(24)00470–2(2024)doi:10.1016/j.ymthe.2024.07.015。8。Anzalone,A。V。等。搜索和重新固定基因组编辑,无需双链断裂或供体DNA。自然576,149–157(2019)。9。Coleman,M。B.等。am。J. Hematol。42,186–190(1993)。 10。 Chen,P。J.等。42,186–190(1993)。10。Chen,P。J.等。Chen,P。J.等。g伽玛A伽马(β+)胎儿血红蛋白的遗传持久性:g伽玛-158 c-> t在顺式中与-175 t-> c c gamma-lobin基因的突变会导致G Gama-- gamma基因的增加导致G Gama-Globobin的增加。通过操纵细胞决定因素的编辑结果来增强质量编辑系统。Cell 184,5635-5652.E29(2021)。 11。 Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Cell 184,5635-5652.E29(2021)。11。Ravi,N。S.等。 通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。 Elife 11,E65421(2022)。 12。 Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Ravi,N。S.等。通过CRISPR基础编辑来识别新型HPFH样突变,从而提高了胎儿血红蛋白的表达。Elife 11,E65421(2022)。12。Kim,H。K.等。 预测人类细胞中主要编辑指南RNA的效率。 nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。 13。 Nelson,J。W.等。 设计的Pegrnas提高了主要的编辑效率。 NAT生物技术40,402–410(2022)。 14。 Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Kim,H。K.等。预测人类细胞中主要编辑指南RNA的效率。nat Biotechnol(2020)doi:10.1038/s41587-020-0677-y。13。Nelson,J。W.等。设计的Pegrnas提高了主要的编辑效率。NAT生物技术40,402–410(2022)。14。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。 核酸Res 50,1187–1197(2022)。 15。 Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Habib,O.,Habib,G.,Hwang,G.-H。 &Bae,S。人类胚胎干细胞中主要编辑结果的全面分析。核酸Res 50,1187–1197(2022)。15。Lee,J。等。 prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。 nat Commun 14,1786(2023)。 16。 Antoniou,P。等。 基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。 nat Commun 13,6618(2022)。 17。 Pavani,G。等。 通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。 血液Adv 5,1137–1153(2021)。 18。Lee,J。等。prime用真正的cas9 nickases编辑最大程度地减少了不需要的indels。nat Commun 14,1786(2023)。16。Antoniou,P。等。基础编辑介导的γ-球蛋白顺式调节元件的解剖,用于胎儿血红蛋白表达的治疗重新激活。nat Commun 13,6618(2022)。17。Pavani,G。等。通过人类造血干细胞中α-珠蛋白基因座的CRISPR/CAS9编辑对β-丘脑的抗性。血液Adv 5,1137–1153(2021)。18。Everette,K。A.等。在体内造血干细胞的体内质量编辑促进小鼠植入后镰状细胞疾病表型。nat Biomed Eng 7,616–628(2023)。19。Peterka,M。等。利用DSB修复以促进有效的同源性依赖性和 - 独立的质量编辑。nat Commun 13,1240(2022)。20。Magnani,A。等。对镰状细胞疾病的同种异体移植后混合嵌合体患者进行了广泛的多系数分析:对基因治疗的造血和植入阈值的见解。Haematologica 105,1240–1247(2020)。21。Sun,Y。等。 在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Sun,Y。等。在小鼠中耐用基因校正的肺部干细胞的体内编辑。 科学384,1196–1202(2024)。 22。 Doman,J。L.等。 噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。 单元格186,3983-4002.E26(2023)。 23。 Wimberger,S。等。 同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。 nat Commun 14,4761(2023)。 24。 Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。在小鼠中耐用基因校正的肺部干细胞的体内编辑。科学384,1196–1202(2024)。22。Doman,J。L.等。噬菌体辅助进化和蛋白质工程产生紧凑,有效的主要编辑者。单元格186,3983-4002.E26(2023)。23。Wimberger,S。等。同时抑制DNA-PK和POLθ提高了基因组编辑的整合效率和精度。nat Commun 14,4761(2023)。24。Yan,J。等。 用内源性的小RNA结合蛋白改善原始编辑。 自然628,639–647(2024)。 25。 Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。 nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。 26。 核酸res。Yan,J。等。用内源性的小RNA结合蛋白改善原始编辑。自然628,639–647(2024)。25。Levesque,S.,Cosentino,A.,Verma,A.,Genovese,P。&Bauer,D。E.通过调节核苷酸代谢,增强造血干和祖细胞中的质量编辑。nat Biotechnol(2024)doi:10.1038/s41587-024-02266-4。26。核酸res。Brinkman,E。K.,Chen,T.,Amendola,M。&Van Steensel,B。通过序列痕量分解对基因组编辑的易于定量评估。42,E168(2014)。 27。 Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和42,E168(2014)。27。Brusson,M。等。 新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。 mol the核酸32,229–246(2023)。 28。 Gaudelli,N。M.等。 腺嘌呤基础编辑者的定向演变,活动增加和Brusson,M。等。新型的慢病毒载体,用于结合基因添加和基因沉默策略的镰状细胞疾病基因治疗。mol the核酸32,229–246(2023)。28。Gaudelli,N。M.等。腺嘌呤基础编辑者的定向演变,活动增加和