XiaoMi-AI文件搜索系统
World File Search SystemHedgehog

靶向蛋白质降解揭示 BET 溴结构域是 Hedgehog 通路抑制剂-1 的细胞靶标
*通讯地址:sascha.hoogendoorn@unige.ch 摘要 从表型筛选中得到的小分子命中物的靶标反卷积是一项重大挑战。许多筛选都表明,人们已进行许多筛选来寻找 Hedgehog (Hh) 信号通路的抑制剂,Hedgehog (Hh) 信号通路是一条与健康和疾病有着诸多关系的主要发育通路,其中有许多命中物但很少有确定的细胞靶标。我们在此提出一种基于蛋白水解靶向嵌合体 (PROTAC) 结合无标记定量蛋白质组学的靶标识别策略。我们开发了一种基于下游 Hedgehog 通路抑制剂-1 (HPI-1) 的 PROTAC,HPI-1 是一种具有未知细胞靶标的表型筛选命中物。使用我们的 Hedgehog 通路 PROTAC (HPP),我们确定并验证了 BET 溴结构域是 HPI-1 的细胞靶标。此外,我们发现 HPP-9 通过延长 BET 溴结构域降解时间,具有作为长效 Hh 通路抑制剂的独特作用机制。总之,我们提供了一种强大的基于 PROTAC 的靶标反卷积方法,该方法回答了 HPI-1 的细胞靶标这个长期存在的问题,并产生了第一个作用于 Hh 通路的 PROTAC。主要 Hedgehog 通路是一个复杂的细胞信号级联,可调节胚胎发育过程,例如模式化,以及干细胞维持和组织稳态。1,2 Hedgehog 信号转导生理水平的失调会导致发育障碍以及各种癌症的发生和进展,最显著的是基底细胞癌和髓母细胞瘤。3,4 正常条件下的通路激活是由其中一种 Hedgehog 蛋白 (IHH、DHH、SHH) 与受体 Patched (PTCH1) 结合启动的。 5–7 HH 与 PTCH1 结合可释放后者对 Smoothened (SMO) 的抑制作用。8,9 进一步的激活步骤包括与融合抑制因子 (SUFU) 结合的 GLI2/3 转录因子通过初级纤毛的尖端运输并积累。10–13 GLI 转录因子加工成其转录活性形式,然后导致 Hedgehog 靶基因的转录,其中包括正调节剂 Gli1 和负反馈回路中的 Ptch1。14,15 目前,唯一获得临床批准用于对抗 Hh 通路驱动癌症的药物是针对 SMO(vismodegib、sonidegib)的药物。由下游通路激活驱动的癌症本质上对这些药物不敏感,并且最初有反应的肿瘤获得性耐药很常见。16–

绿色氨作为空间能量矢量:评论
摘要:哺乳动物结合的O-Acyltransfer- ASE(MBOAT)超家族参与了生长,发育和食欲感应的生物学过程。MBoats是癌症和肥胖症中有吸引力的药物靶标。但是,关于小分子抑制的结合位点和分子机制的信息是难以捉摸的。这项研究报告了光化学探针的合理发展,以询问人类Mboat Hedgehog酰基转移酶(HHAT)中新型的小分子抑制剂结合位点。结构 - 活性关系研究确定了单个启动IMP-1575,这是最有效的HHAT抑制剂报告的待办事项,并指导了维持HHAT抑制效力的Photocroprosslink探针的设计。光叠链链接和HHAT的蛋白质组学测序对哺乳动物MBoat中的第一个小分子结合位点进行了鉴定。拓扑和同源性数据提出了HHAT抑制的潜在机制,该机制已通过动力学分析证实。我们的结果提供了最佳的HHAT工具抑制剂IMP-1575(K I = 38 nm),并提供了绘制MBOAT中小分子相互作用位点的策略。m的膜结合的O-酰基转移酶(MBOAT)蛋白质的超家族与几种至关重要的生物学途径有关。[1]在人类中,其中包括Wnt酰基转移酶(豪猪; Porcn),[2] Hedge- Hog酰基转移酶(HHAT)[3]和Ghrelin O -acyltransferase(Goat)[4],这些酶(山羊)[4]分别调节Wnt和HedgeHog信号传播以及食欲。这些MBoats是癌症和肥胖症中有吸引力的治疗靶标,[1],哺乳动物MBoats的结构信息受到了极大的追捧。各种哺乳动物MBoats的膜拓扑已通过实验确定,支持保守的

vemurafenib激活Sonic刺猬途径并促进甲状腺癌干细胞自我更新
B-RAF激酶抑制剂,例如vemurafenib(PLX4032)和dabrafenib对BRAF氧化甲状腺癌的治疗功效有限。癌症干细胞(CSC)在肿瘤复发,耐药性和转移中起重要作用。CSC是否在抑制B-RAF激酶抑制剂的抗肿瘤活性中发挥作用仍然未知。在这里,我们报告说,vemurafenib(PLX4032)在两种间隙性甲状腺癌细胞系中引起了几种相关基因的表达,包括Gli1,Snail,BMI1和Sox2,SW1736和8505C,但在A375细胞中降低了A375细胞的表达,A375细胞的表达,A375细胞中,A375细胞的表达。PLX4032促进了甲状腺癌干细胞自我更新,这是醛醛脱氢酶阳性细胞和甲状腺酸盐数量增加所证明的。从机械上讲,PLX4032激活了通过HER3激活PI-3和有丝分裂原激活的蛋白激酶途径,以交叉激活Gli1(Sonic Hedgehog(SHH)途径的转录因子)。gant61是一种特异性GLI1的抑制剂,阻止了在PLX4032处理的甲状腺癌细胞的体外和体内在两种甲状腺癌异种移植模型中的表达。gant61仅处理弱抑制了SW1736肿瘤的生长,但在组合使用时会增强PLX4032的抗肿瘤活性。我们的研究提供了有关甲状腺癌如何对B-RAF激酶抑制剂反应较差的机械见解,并表明将B-RAF和SHH途径组合起来可能会克服甲状腺癌耐药性。

了解 Hedgehog 信号通路在 T 细胞淋巴细胞生成和 T 细胞急性淋巴细胞白血病 (T-ALL) 中的作用
摘要:Hedgehog (HH) 信号网络是无脊椎动物和脊椎动物胚胎发育的主要调节器之一。与其他网络(如 NOTCH 和 WNT)一起,HH 信号通过时间和空间调节细胞增殖和分化来指定早期模式和极性事件以及随后的器官形成。然而,已在多种恶性疾病中发现 HH 信号的异常激活,它对肿瘤细胞的增殖、存活和治疗耐药性有积极影响。针对 HH 通路的抑制剂已在临床前癌症模型中进行了测试。HH 通路在其他血液恶性肿瘤中也过度活跃,包括 T 细胞急性淋巴细胞白血病 (T-ALL)。本综述旨在总结我们对正常 T 细胞淋巴细胞生成和 T-ALL 中 HH 通路的生物学作用和病理生理学的了解。此外,我们将讨论可能扩大针对 T-ALL 中的 HH 通路的药物临床用途的潜在治疗策略。

刺猬途径抑制剂的局部晚期和转移性基底细胞癌:现实世界中的单中心回顾性评论
基底细胞癌(BCC)可以通过手术切除或辐射高度治愈。在极少数情况下,BCC可能是局部破坏性的,也可能难以手术去除。用Vismodegib或Sonidegib的刺猬抑制(HHI)诱导50–60%的响应率。长期毒性包括肌肉痉挛和体重减轻,导致剂量降低。此回顾性图表审查还研究了COQ10和补充钙补充对HHI药物在2012年至2022年接受HHI药物治疗的患者的影响。我们审查了用Vismode-Gib或Sonidegib治疗的局部晚期或转移性BCC的成年患者的图表,主要用于无进展生存率(PFS)。次要目标包括总体生存,特定于BCC的生存,停止的时间和原因,总回应率,安全性和耐受性,COQ10和钙补充剂的使用以及保险范围。在55例可评估结果的患者中,有34例(61.8%)具有整体临床益处,有25例(45.4%)具有完全反应,而9(16.3%)的部分反应。在14(25.4%)和7(12.7%)中发现了稳定的疾病。 对治疗反应的34例患者有9例重复。 被HHI重新收录的患者可以再次反应。 5年(5年)总体BCC特异性生存率的中位数为89%。 Vismodegib和Sonidegib的剂量减少或终止,分别为59%,案例为24%,分别为30%和9%的病例。 具有COQ10和补充钙的补充,只有17%的剂量减少,而没有剂量的42%。在14(25.4%)和7(12.7%)中发现了稳定的疾病。对治疗反应的34例患者有9例重复。被HHI重新收录的患者可以再次反应。5年(5年)总体BCC特异性生存率的中位数为89%。Vismodegib和Sonidegib的剂量减少或终止,分别为59%,案例为24%,分别为30%和9%的病例。具有COQ10和补充钙的补充,只有17%的剂量减少,而没有剂量的42%。HHI对于治疗晚期BCC非常有效,但可能需要减少给药。sonidegib比Vis-Modegib更好地容忍。COQ10和补充钙可以有效预防肌肉痉挛。COQ10和补充钙可以有效预防肌肉痉挛。

刺猬激活通过瞬态克隆能力
简介生长板 - 位于长骨边缘的薄盘状软骨 - 为产后骨骼生长提供了主要驱动力(1)。从结构上讲,生长板由3个形态学 - 静止,增殖和肥厚的区域组成,具有具有克隆起源的软骨细胞的特征柱(2,3)。生长板是内侧软骨骨形成的必不可少的结构,该过程逐渐被骨骼逐渐取代(1,2)。位于产后生长板顶部的静息区载有慢循环软骨细胞,表达甲状旁腺激素相关蛋白(PTHRP)(4),该蛋白(4)提供了生长板中所有其他软骨细胞的来源。这些“静止”的软骨细胞通过不对称分裂进入细胞周期,成为增殖的软骨细胞,分化为表达印度刺猬(IHH)的有丝虫后自生型前软骨细胞(IHH),变成肥大的软骨细胞的生长板和死亡的骨骼或因素而变成骨的底部或因素而变成骨的底部或因素而转变为骨的底部,因为主要海绵中的成骨细胞。

克服对刺猬抑制剂的耐药性:同时发生的Sonidegib和放射疗法的第一个临床病例,用于在刺猬抑制剂上进展的晚期基底细胞癌
这位患者出生于1933年,是一名90岁的女性,具有过去的动脉高血压病史。她的癌症史始于鼻尖皮肤病变,随后她是皮肤科医生,并进行了局部治疗。两年后,病变开始迅速生长,伴有右鼻子沟的潜在肿胀,骨骼变形和鼻金字塔的大溃疡。核心活检于2021年9月进行,显示基底细胞癌(结节性)。该患者去了另一家医院,并被一位新的皮肤科医生拜访,该医生报告了涉及右Alar缘,鼻尖和右鼻腔侧壁的3厘米病变的存在。鉴于在没有整容畸形的情况下对手术切除的广泛疾病过敏,她被认为不适合手术,被称为医学肿瘤学。2021年9月7日,她被医学肿瘤学家看到,并开始使用Sonidegib(每天200毫克)(图1和2; 2021年9月2日)。她的最初部分反应,BCC大小的临床减少(图3; 2022年8月),除了1级疲劳和1级dydysgeusia外,副作用最小。部分缓解一直持续到2022年12月,没有明显的毒性,然后自2023年1月以来就在临床上进行。

在神经细胞原发性纤毛的钙动力学调节刺猬信号传导依赖性神经发生在胚胎神经管
神经干细胞增殖与神经元分化之间的平衡对于适当发展神经系统至关重要。Sonic刺猬(SHH)依次促进细胞增殖和神经表型的规范,但是负责从有丝分裂到神经源的发育转变的信号传导机制尚不清楚。在这里,我们表明,SHH通过瞬态受体电势阳离子阳离子c构件C成员3(TRPC3)(TRPC3)通过Ca 2+涌入来增强Ca 2+的活性,并通过Ca 2+涌入发育于Ca 2+涌入,并以发育阶段相互依赖的阶段相关的方式从细胞内存储中释放。这种睫状Ca 2+的活性反过来又通过下调SOX2表达和神经源性基因的上调表达来拮抗神经干细胞中的规范,增生性SHH信号,从而实现了神经元分化。这些发现表明,神经细胞睫状信号传导中的SHH-CA 2+依赖性开关触发了SHH作用从规范有限源性到神经源的开关。在该神经源信号轴上鉴定的分子机制是治疗脑肿瘤和神经发育障碍的潜在靶标。
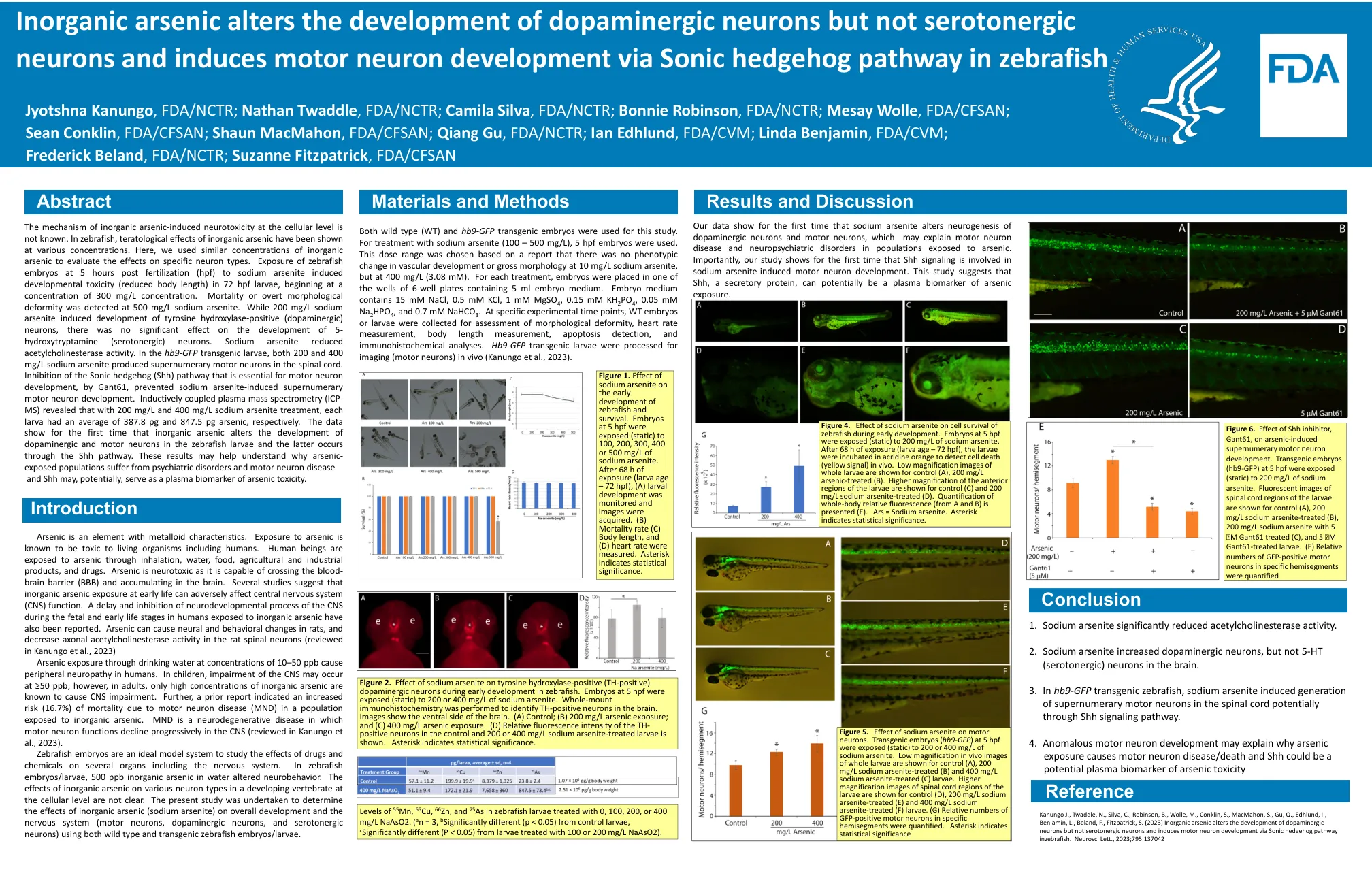
无机砷改变多巴胺能神经元的发育,但不改变血清素能神经元的发育,并通过 Sonic Hedgehog 通路诱导斑马鱼的运动神经元发育
无机砷在细胞水平上诱发神经毒性的机制尚不清楚。在斑马鱼中,不同浓度的无机砷均有致畸作用。在这里,我们使用了类似浓度的无机砷来评估其对特定神经元类型的影响。受精后 5 小时 (hpf) 的斑马鱼胚胎暴露于亚砷酸钠中,在 72 hpf 幼虫中诱发发育毒性(体长缩短),浓度从 300 mg/L 开始。在 500 mg/L 亚砷酸钠下检测到死亡或明显的形态畸形。虽然 200 mg/L 亚砷酸钠诱导酪氨酸羟化酶阳性(多巴胺能)神经元的发育,但对 5-羟色胺(血清素能)神经元的发育没有显著影响。亚砷酸钠降低了乙酰胆碱酯酶活性。在hb9-GFP转基因幼鱼中,200和400mg/L亚砷酸钠均在脊髓中产生了多余的运动神经元。通过Gant61抑制运动神经元发育所必需的Sonic Hedgehog(Shh)通路,可以阻止亚砷酸钠诱导的多余运动神经元发育。电感耦合等离子体质谱(ICP-MS)分析表明,在200mg/L和400mg/L亚砷酸钠处理下,每只幼鱼平均砷含量分别为387.8pg和847.5pg。数据首次表明无机砷改变斑马鱼幼鱼多巴胺能神经元和运动神经元的发育,后者是通过Shh通路发生的。这些结果可能有助于理解为什么接触砷的人群会患上精神疾病和运动神经元疾病,并且 Shh 可能潜在地充当砷毒性的血浆生物标志物。

env-101是一种新型的刺猬抑制剂,可提高肺功能并降低特发性肺纤维化患者的肺纤维化:由A
缩写:AE,不利事件;阿拉特,拉丁美洲胸部协会; ATS,美国胸腔学会; BMI,体重指数; CT,计算机断层扫描; DL CO,肺部碳一氧化碳的扩散能力;心电图,心电图; ERS,欧洲呼吸社会; FVC,强迫生命力; HRCT,高分辨率CT; IPF,特发性肺纤维化; JRS,日本呼吸社会; PBO,安慰剂; PO,口头; PPF,进行性肺纤维化; QD,每天一次; QGG,定量地面玻璃; QILD,定量间质肺疾病; QLF,定量肺纤维化; SAE,严重的不利事件; SD,标准偏差;嘘,声音刺猬; Smo,平滑; TEAE,治疗急性不良事件; TLC,总肺容量;加利福尼亚大学圣地亚哥分校; W12,第12周。