XiaoMi-AI文件搜索系统
World File Search SystemLACTAM

青霉素* 类别:β-内酰胺概述
青霉素* 类别:β-内酰胺 概述 青霉素是典型的β-内酰胺,至今仍是一种重要的抗菌药物。兽医学中使用的窄谱青霉素包括天然存在的青霉素、青霉素 G (苄基青霉素) 和口服的生物合成稳定青霉素,如青霉素 V (苯氧甲基青霉素) 和苯乙青霉素。青霉素对大多数敏感细菌有杀菌作用。这些窄谱 β - 内酰胺酶敏感青霉素主要对抗革兰氏阳性菌。 耐药性 在 20 世纪 40 年代初青霉素广泛使用后不久,产生青霉素酶的金黄色葡萄球菌开始出现。青霉素酶通过酶抑制破坏β - 内酰胺环,使菌体产生耐药性。耐药性的垂直进化是指通过突变和选择获得耐药性。通过这一过程,肺炎球菌能够改变青霉素结合的靶蛋白。先前对青霉素敏感的生物体也可以通过转导获得耐药性。耐药金黄色葡萄球菌可以通过将包裹在噬菌体、细菌宿主病毒中的质粒 DNA 转移给敏感金黄色葡萄球菌,从而将 β-内酰胺酶产生能力转移给敏感金黄色葡萄球菌。生物体还可以通过转化、从死细菌中吸收暴露的 DNA 并导致基因型改变(DNA 重组)获得耐药性。这可能是肺炎链球菌传播青霉素耐药性的主要手段。结合是获得青霉素耐药性的另一种方式。结合涉及同属或不同属的细菌之间质粒或其他染色体外 DNA 的单向转移。这种转移通过生育因素介导发生,并通过性菌毛从供体延伸到受体进行。这种机制通常是多药耐药性转移的原因。这种现象发生在一系列携带抗多种抗菌药物的耐药性转移因子的紧密相关基因发生交换时。这些质粒通过转移含有形成 β -内酰胺酶所需信息的基因来携带对青霉素的耐药性。

氨苄西林* 类别:β-内酰胺概述
氨苄西林* 类别:β-内酰胺 概述 氨苄西林,俗称广谱青霉素,是一种氨基青霉素,是一类半合成的 β-内酰胺,专门用于对抗革兰氏阴性菌和革兰氏阳性菌。氨基青霉素是通过将青霉素与氨基或侧链连接而生成的。添加侧链会显著改变药物对某些细菌的活性。最初,这些抗菌药物对奇异变形杆菌、大肠杆菌、志贺氏菌、沙门氏菌、嗜血杆菌和奈瑟菌有效。然而由于易感性的变化,氨苄西林不再是治疗这些菌感染(如大肠杆菌尿路感染)的首选药物,除非培养和药敏结果表明易感。氨苄西林的作用机制是通过附着于青霉素结合蛋白 (PBP) 来干扰细胞壁合成,抑制细胞壁肽聚糖合成和使自溶酶抑制剂失活。耐药性 氨苄西林通常也被 β-内酰胺酶灭活(有关获得对 β-内酰胺的耐药性的信息,请参阅青霉素部分。)。近年来,屎肠球菌和肺炎链球菌开始通过突变表现出低亲和力 PBP,这是对氨基青霉素的耐药机制。有效性 氨苄西林和阿莫西林具有相同的活性谱,尽管阿莫西林的特点是生物利用度更高。对氨苄西林和阿莫西林普遍敏感的菌属包括葡萄球菌、链球菌、棒状杆菌、梭状芽孢杆菌、大肠杆菌、克雷伯氏菌、志贺氏菌、沙门氏菌、变形杆菌和巴氏杆菌,尽管其中许多细菌已获得耐药性。氨苄西林通常用于治疗革兰氏阴性肠道细菌引起的泌尿道感染。该药物还用于治疗呼吸道感染。此外,氨苄西林对 B 组链球菌均有效,但对肠杆菌、流感嗜血杆菌、假单胞菌和吲哚阳性变形杆菌感染无效。有关体液和脑脊液吸收的解释,请参阅青霉素部分。 *可根据要求提供参考资料。致电路易斯安那州卫生与医院部公共卫生办公室传染病流行病学科 (504-219-4563)

β-内酰胺抗生素可以阻止体液免疫吗?
据报道,用B -LACTAM抗生素治疗诱导白细胞减少症和念珠菌病,使对抗癌免疫疗法的临床反应恶化,并降低对疫苗接种的免疫反应。b-乳糖酶可以通过阻断其活性来切割B-乳酰胺抗生素。描述了B-乳糖苷酶的两个不同的超家族,丝氨酸B-乳糖苷酶和锌离子依赖的金属B-乳酰胺酶。在人类中,已经鉴定出了18个金属B-乳糖酶编码基因(HMBL)。虽然大多数人的生理作用仍然未知,但已经很好地确定SNM1A,B和C蛋白参与DNA修复。SNM1C/Artemis蛋白在V(d)J段重排中完全相关,这会导致免疫球蛋白(Ig)和T细胞受体变量区域,在免疫反应中具有至关重要的作用。因此,在人类中,SNM1C/Artemis突变与严重的合并免疫差异有关,其特征在于低磁性血症有效的细胞免疫和机会性感染。虽然HMBL的催化位点,尤其是SNM1家族的催化位点是高度保守的,但体外研究表明,一些B- lactam抗生素以及精确的第三代头孢菌素和氨苄青霉素抑制了金属 - B-乳糖酶蛋白SNM1A&B和SNM1C/ARTM1C/ARTEMIS蛋白质。类比,出现了一个问题,即B-乳酰胺抗生素是否可以阻止人类诱导短暂免疫降低的人类SNM1C/Artemis蛋白。我们在这里审查了基于计算机,体外和体内证据的基于该假设的文献数据。了解B -LACTAM抗生素对免疫细胞的影响将提供新的治疗线索和肿瘤学,免疫学和传染病的新临床方法。
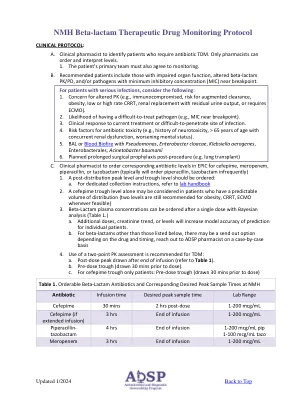
NMHβ-LACTAM TDM协议
f t>麦克风地板/病房100%> 1x MIC所有缩写:ICU,重症监护室; MIC,最小抑制浓度; SCT,干细胞移植; SOT,固体器官移植 *应根据患者的临床状况选择PK/PD靶标的重病患者以及患有深度座椅感染或免疫功能低下的状态的患者的临床状况,请考虑选择患者对治疗疗法的状态和反应的更具侵略性的PK/PD靶标。

遗传决定因素是大肠杆菌中β-内酰胺/β-内酰胺酶抑制剂耐药性的进行性表型的遗传决定因素
此预印本的版权持有人(本版本发布于2023年5月25日。; https://doi.org/10.1101/2023.05.24.542208 doi:Biorxiv Preprint

开发靶向PBP2A的非β-内酰胺共价抑制剂在耐甲氧西林的金黄色葡萄球菌
耐甲氧西林金黄色葡萄球菌(MRSA)是一种革兰氏阳性的细菌病原体,继续对我们社会中当前的公共卫生系统构成严重威胁。MRSA中对β-内酰胺抗生素的高度抗性归因于青霉素结合蛋白2a(PBP2A)的表达,这会催化细胞壁交联。根据大量研究报告,已知PBP2A蛋白的活性受到与细胞壁交联的活性位点不同的变构位点的调节。在这里,我们对包含1,3,4-氧化唑核的113种化合物进行了筛选,以设计针对PBP2A变构位点的新共价抑制剂并建立其结构活性关系。在初始筛选中鉴定出的磺酰氧化二唑化合物的立体选择性合成导致细胞抑制活性的最大增强。基于基于PEG的药膏的磺酰基黄烷二唑的化合物,对人细胞的毒性测试低(CC 50:>78μm),不仅在小鼠皮肤伤口感染模型中,而且还针对抗氧蛋白抗抗性临床分离型MRSA(IC 50ous)(IC 50oubious),表现出了有效的抗菌作用。此外,利用LC-MS/MS和硅内方法的其他研究清楚地支持了通过亲核芳香族反应(S NAR)反应(S NAR)的变构位点共价结合机制,以及与PBP2A主要活性位点关闭的关联。

- 乳糖酶抑制剂作为抗菌化疗中的增强剂的作用,靶向革兰氏阴性细菌
摘要:由于发现青霉素,β -lactam抗生素通常用于治疗细菌感染。不幸的是,与此同时,病原体可以通过产生β-乳糖酶来发展对β-乳酰胺抗生素的抗性,例如青霉素,头孢菌素,单oc省和碳青霉烯。因此,将β -LACTAM抗生素与β-内酰胺酶抑制剂的组合是控制β-lactActam抗性细菌的一种有希望的方法。新型β-乳糖酶抑制剂(BLI)的发现对于有效治疗抗生素耐药细菌感染至关重要。因此,这篇综述讨论了旨在增强β-lactam抗生素活性的创新抑制剂的发展。具体而言,本综述描述了不同类别的β-乳糖酶的分类和特征以及β-乳酰胺和BLI的协同机制。此外,我们还引入了化合物的潜在来源,以用作新型BLIS。这为克服β-乳糖果酶产生细菌的当前挑战提供了见解,并与BLI结合设计有效的治疗选择。

![vabomere - [产品专论模板 - 标准]](/simg/b\bc185d48caf196f4f8a9d8111f5fc6381befee8a.webp)
vabomere - [产品专论模板 - 标准]
接受 β-内酰胺类抗生素(包括注射用美罗培南)治疗的患者中,曾报告出现严重且偶尔致命的超敏反应。这些反应更可能发生在对多种过敏原有过敏史的个体身上(见 8.5 上市后不良反应)。有报告称,有青霉素超敏反应史的个体在接受其他 β-内酰胺类抗生素治疗时出现严重反应。在开始注射用美罗培南治疗之前,应仔细询问之前对青霉素、头孢菌素、其他 β-内酰胺类抗生素和其他过敏原的超敏反应。如果对注射用美罗培南出现过敏反应,应立即停用该药。过敏反应需要立即用肾上腺素治疗。可能需要氧气、静脉注射类固醇、抗组胺药和气道管理,包括插管。