XiaoMi-AI文件搜索系统
World File Search SystemLinker
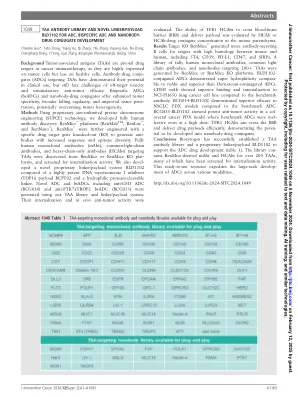
背景 肿瘤相关抗原(TAA)是癌症免疫治疗中的关键药物靶点,因为它们在肿瘤细胞上表达较高,而在健康细胞上表达较少。靶向 TAA 的抗体-药物偶联物(ADC)已证明其在临床应用中的潜力,但仍然面临着脱靶毒性和抗肿瘤疗效不理想的挑战。双特异性 ADC(bsADC)和纳米抗体-药物偶联物具有增强的肿瘤特异性、更广泛的杀伤能力和更好的组织穿透性,有可能克服肿瘤异质性。方法 利用专有的无限大小精准染色体工程(SUPCE)技术,我们开发了完全人源抗体发现 RenMice ® 平台(RenMab TM 、RenLite ® 和 RenNano ® )。RenMice ® 经过进一步工程改造,具有特定的药物靶标基因敲除(KO),以产生具有增加的序列和表位多样性的抗体。从 RenMice 或 RenMice KO 平台发现了靶向 TAA 的全人源单克隆抗体 (mAb)、共同轻链抗体和重链抗体 (HCAb),并筛选了其内化活性。我们还开发了一种新型专有接头/有效载荷系统 BLD1102,该系统由高效 DNA 拓扑异构酶 I 抑制剂 (TOP1i) 有效载荷 BCPT02 和亲水性蛋白酶可裂解接头组成。使用我们的 TAA 库和接头/有效载荷系统生成了新型 ADC 和 bsADC,包括抗 CH3 ADC (BCG014) 和抗 PTK7xTROP2 bsADC (BCG033)。它们的内化和体内抗肿瘤活性

分子对接,MD模拟,合成,DFT和生物学评估以及与Para Nitro苯基
伏诺替纳斯特的结构特征显示了三个部分,例如表面识别苯甲酰胺,接头己酰基和金属结合羟氨酸。在这项工作中,用取代的苯基环改变了表面识别组,咪唑基 - 三唑组用相同的金属结合羟氨基酸更改了接头组,最后设计了(F1-F4)分子。然后将所有设计的分子对接使用HDAC 2(4LXZ)受体。f4显示-8.7 kcal/mol的最大结合能,标准vornostat显示-7.2 kcal/mol。所有设计的分子都是使用gromacs软件模拟的分子动力学,以确定RMSD,RMSF,SASA和氢键的数量。所有仿真数据显示配体和受体之间的良好相互作用。然后,所有分子均由三个部分合成:a。二硝基苯基连接的三唑羟酸的合成,b。取代的恶唑酮衍生物的合成和c。在最后一步中,对替代的恶唑酮衍生物和二硝基苯基链接的三唑羟氨基酸反应,以产生最终的分子集(F1-F4)。DFT分析确定,F4以良好的亲电性而出现为最反应性分子。此外,对乳腺癌细胞系的体外抗增殖活性表明,F4是所有合成分子中最有效的抗癌分子。

AZD8205(一种抗B7-H4抗体 - 药物结合物)广泛的生物转化分析,阐明了其在体内稳定性的途径
体内大分子会发生什么?是什么驱动抗体 - 药物缀合物(ADC)的结构活性关系和体内稳定性?这些相互关联的问题越来越相关,因为ADC作为有影响力的治疗方式的重新重要性以及我们对ADC结构决定因素的理解中存在的差距,而ADC是体内稳定性的ADC结构决定因素。复杂的大分子(例如ADC)可能会因其复杂的结构而发生变化,因为它们可能会在接头,有效载荷和/或在修饰的共轭位点上发生生物转化。此外,由于难以识别或量化大型大分子上的较小变化,ADC代谢的解剖提出了重大的分析挑战。我们采用了免疫接触LCMS方法来评估四种不同铅ADC中药物抗体比(DAR)谱的体内变化。这种全面的特征表明,随着互联网的选择,有助于ADC设计的关键结构决定因素是选择接头,因为复古 - 米克尔脱糖与硫二酰亚胺的水解反应之间的竞争导致体内出色的共轭稳定性。这些数据与其他因素结合了其他因素,告知AZD8205,B7-H4指导的半胱氨酸结合的ADC,带有新型的拓扑异构酶I抑制剂有效载荷,并具有耐用的DAR,目前正在临床上研究固体恶性肿瘤(NCT051223482)。这些结果突出了研究大分子生物转化并阐明ADC结构 - 体内稳定性关系的相关性。这项工作的全面性质增加了对我们的

实时监测爪蟾鸡蛋提取物中单个DNA分子的压实
DNA压实是在有丝分裂过程中凝结和分辨率的凝结和分辨率所必需的,但是单个染色质因子对该过程的相对贡献知之甚少。我们使用高速爪蟾卵提取物和光学镊子开发了一种生理,无细胞的系统,以研究实时有丝分裂染色质纤维的形成,并在单个DNA分子上进行力诱导的拆卸。与将DNA压缩约60%的相间提取物相比,中期提取物将DNA的长度降低了90%以上,这反映了这两种情况下全染色体形态的差异。抑制核小体组装的核心组蛋白伴侣ASF1的耗竭,将中期纤维压实的最终程度降低了29%,而接头组蛋白H1的耗竭效果更大,将总压实降低了40%。 与对照组相比,两种耗竭都降低了压实率,导致了更短的分解时间,并提高了力诱导的纤维拆卸速度。 相比之下,中期提取物中冷凝蛋白的耗竭强烈抑制纤维组件,从而导致瞬态压实事件在高力下迅速逆转。 总的来说,这些发现支持了一种投机模型,在该模型中,冷凝蛋白在有丝分裂DNA压实中起主要作用,而核心和接头组蛋白起作用,可在循环挤出过程中减少滑移并调节DNA压实程度。抑制核小体组装的核心组蛋白伴侣ASF1的耗竭,将中期纤维压实的最终程度降低了29%,而接头组蛋白H1的耗竭效果更大,将总压实降低了40%。与对照组相比,两种耗竭都降低了压实率,导致了更短的分解时间,并提高了力诱导的纤维拆卸速度。相比之下,中期提取物中冷凝蛋白的耗竭强烈抑制纤维组件,从而导致瞬态压实事件在高力下迅速逆转。总的来说,这些发现支持了一种投机模型,在该模型中,冷凝蛋白在有丝分裂DNA压实中起主要作用,而核心和接头组蛋白起作用,可在循环挤出过程中减少滑移并调节DNA压实程度。

定向域间运动使 IsdH 金黄色葡萄球菌受体能够从人体血红蛋白中快速提取血红素
致病性金黄色葡萄球菌利用 IsdH 表面受体主动从人类血红蛋白 (Hb) 中获取铁。血红素提取由受体内的三域单元介导,该单元包含其第二 (N2) 和第三 (N3) NEAT 域,由螺旋连接域连接。提取发生在动态复合体中,其中受体与每个珠蛋白链结合;N2 域与 Hb 紧密结合,而受体内大量的域间运动使其 N3 域能够暂时扭曲珠蛋白的血红素口袋。使用分子模拟结合马尔可夫模型,以及停流实验定量测量血红素转移动力学,我们表明受体内的定向域间运动在提取过程中起着关键作用。N3 域运动的方向性和血红素提取的速率由连接 N2 和连接域的短而灵活的域间系绳内的氨基酸控制。在野生型受体中,源自系链的定向运动使 N3 域能够填充能够扭曲 Hb 口袋的配置,而含有改变的系链的突变受体不太能够采用这些构象异构体并通过间接过程缓慢捕获血红素,其中 Hb 首先将血红素释放到溶剂中。因此,我们的结果表明 IsdH 受体内的域间运动在其能力中起着关键作用

定向域间运动使 IsdH 金黄色葡萄球菌受体能够从人体血红蛋白中快速提取血红素
致病性金黄色葡萄球菌利用 IsdH 表面受体主动从人类血红蛋白 (Hb) 中获取铁。血红素提取由受体内的三域单元介导,该单元包含其第二 (N2) 和第三 (N3) NEAT 域,由螺旋连接域连接。提取发生在动态复合体中,其中受体与每个珠蛋白链结合;N2 域与 Hb 紧密结合,而受体内大量的域间运动使其 N3 域能够暂时扭曲珠蛋白的血红素口袋。使用分子模拟结合马尔可夫模型,以及停流实验来定量测量血红素转移动力学,我们表明受体内的定向域间运动在提取过程中起着关键作用。N3 域运动的方向性和血红素提取的速率由连接 N2 和连接域的短而灵活的域间系绳内的氨基酸控制。在野生型受体中,源自系链的定向运动使 N3 域能够填充能够扭曲 Hb 口袋的配置,而含有改变的系链的突变受体不太能够采用这些构象并通过间接过程缓慢捕获血红素,其中 Hb 首先将血红素释放到溶剂中。因此,我们的结果表明 IsdH 受体内的域间运动在其能力中起着关键作用

抗体-药物偶联物的战略作用
此外,连接技术正在提高键稳定性,防止细胞毒性有效载荷的过早释放,从而最大限度地减少脱靶效应并增强有效载荷向肿瘤细胞的输送。5 具体而言,位点特异性结合方法的改进被用于生产具有一致药物抗体 (DAR) 比率的均质 ADC,从而改善治疗和药代动力学指标。6 除了对单个组件进行工程设计外,研究人员还在寻求重新定义 ADC 策略,同时探索双特异性抗体、双有效载荷和非内化抗体等双靶向方法,以克服耐药性并提高特异性。

抗体 - 药物结合物的战略作用
此外,接头技术正在改善键稳定性,以防止细胞毒性有效载荷的过早释放,从而最大程度地降低脱靶效应并增强有效载荷向肿瘤细胞的传递。5具体,正在利用改进位点特异性共轭方法的改进,以产生具有一致的药物对抗体(DAR)比率的同质ADC,从而改善了治疗和药代动力学指数。6除了单个组件的工程外,研究人员还试图通过探索双重目标方法来重新定义ADC策略,例如双特异性抗体,双付费载荷和非内在化抗体来克服耐药性和增强特异性。

抗体 - 药物结合物的战略作用
此外,接头技术正在改善键稳定性,以防止细胞毒性有效载荷的过早释放,从而最大程度地降低脱靶效应并增强有效载荷向肿瘤细胞的传递。5特别是,正在利用特定地点结合方法的改进来生产具有一致的药物与抗体(DAR)比率的均质ADC,从而改善了治疗和药代动力学指数。6除了单个组件的工程外,研究人员还试图通过探索双重目标方法来重新定义ADC策略,例如双特异性抗体,双付费载荷和非内在化抗体来克服耐药性和增强特异性。

利用 TadA 的下一代胞嘧啶碱基编辑器
• 脱氨酶的定向进化 • PAM 变体碱基编辑器 • 定向进化 Cas9 以创建用于 BE 的非 NGG PAM 变体 • 密码子、NLS 和接头优化 • 环状置换体和镶嵌碱基编辑器 • DNA 脱靶评估 • RNA 脱靶评估 • 旁观者编辑最小化 • 引导 RNA 工程 • 离体和体内 BE 递送 • 最小化脱靶活性的工程 BE • HSC、肝细胞和 T 细胞的离体碱基编辑 • ABE 的低温电子显微镜结构 • 小鼠体内碱基编辑 • 非人类灵长类动物体内编辑