XiaoMi-AI文件搜索系统
World File Search SystemMDA5

MDA5依赖性响应 - to-tribute-to-AutoMune- ...
引言1型糖尿病(T1D)是T细胞介导的自身免疫性疾病,导致胰腺β细胞破坏(1)。提出了遗传学,环境和免疫系统的协同作用来诱导T1d(2-5)。单子双胞胎的T1D具有约30%–50%的一致性,这表明环境在T1D发育中起着重要作用(6,7)。与T1D相关的一个环境因素是Coxsackievivirus B(CVB)感染(8,9)。CVB病毒RNA和/或病毒颗粒已在最近发作T1D的患者的血液,粪便和胰岛中检测到(9-11)。在非肥胖糖尿病(NOD)小鼠模型中,CVB感染通过诱导炎症性胰腺抗病毒反应加速T1D,导致β细胞破坏(12,13)。由IFIH1基因编码的先天病毒传感器分化相关蛋白5(MDA5)检测到DSRNA病毒复制中间体并启动抗病毒信号传导(14,15)。MDA5结合其配体后的关键反应之一是I型IFN的合成,例如IFN-α和IFN-β,以促进巨噬细胞,树突状细胞和T细胞的病毒清除和激活(16-20)。尽管I型IFN对抗病毒反应至关重要,但它们也与早期T1D发育有关(21,22)。在转基因CD1小鼠中,其中β细胞组成型表达IFN-α,T1D发作发生在10周龄的60%小鼠中(23)。相反,在NOD雌性小鼠中,IFN -α和-β受体亚基1(IFNAR1)表达的丧失导致T1D发育显着延迟(24)。在T1D患者中,在自身抗体发育之前在血液中检测到I型IFN基因特征(21、22),而GWAS发现与I型IFN合成和信号传导有关的T1D基因,例如IFIH1(例如IFIH1)(25,26)。IFIH1中的多个单核苷酸多态性(SNP)与人类T1D发育有关。A946T SNP(rs1990760)在氨基酸946时导致丙氨酸对硫代的变化,与T1D风险相关,并导致IFN-α /β和IFN刺激的基因产生< / div>的增加

RNA 编辑在有限数量的位点足以阻止小鼠大脑中的 MDA5 激活
作用于 RNA 1 的腺苷脱氨酶 (ADAR1) 是一种负责腺苷到肌苷 RNA 编辑的酶,由两种亚型组成:核 p110 和细胞质 p150。小鼠中 Adar1 或 Adar1 p150 基因的缺失会导致胚胎致死,并伴有干扰素刺激基因 (ISG) 的过表达,这是由于黑色素瘤分化相关蛋白 5 (MDA5) 对未编辑的内源转录本的异常识别所致。然而,在众多 RNA 编辑位点中,有多少 RNA 位点需要编辑,尤其是由 ADAR1 p150 编辑,以避免 MDA5 激活,以及 ADAR1 p110 是否有助于此功能仍不清楚。具体来说,ADAR1 p110 在小鼠脑中含量丰富,而 ADAR1 p150 的表达量微乎其微,而 ADAR1 突变会导致艾卡迪-痛风综合征,在这种综合征中,大脑是受影响最严重的器官之一,同时伴有 ISG 表达升高。因此,了解 RNA 编辑介导的预防大脑中 MDA5 活化的方法尤为重要。在这里,我们建立了 Adar1 p110 特异性敲除小鼠,在这种小鼠中未观察到 ISG 表达上调。这一结果表明 ADAR1 p150 介导的 RNA 编辑足以抑制 MDA5 活化。因此,我们进一步创建了 Adar1 p110 / Adar2 双敲除小鼠来确定 ADAR1 p150 介导的编辑位点。这项分析表明,尽管没有观察到 ISG 表达升高,但在 Adar1 p110 / Adar2 双敲除小鼠的大脑中,只有不到 2% 的编辑位点得以保留。值得注意的是,我们发现一些位点被高度编辑,与野生型小鼠的编辑位点相当,这表明存在 ADAR1 p150 特异性位点。这些数据表明,在非常有限的位点上进行 RNA 编辑(由少量 ADAR1 p150 介导)足以阻止 MDA5 激活,至少在小鼠大脑中是如此。

结核分枝杆菌激活 MDA5 RNA 传感通路促进先天免疫颠覆和病原体存活
引言结核病 (TB) 仍然是一项严重的健康挑战,仅在 2021 年全球就造成约 150 万人死亡 (1)。结核分枝杆菌 (M . tuberculosis) 具有极强的人类适应性,通过尚不完全了解的免疫破坏机制在巨噬细胞内存活。肺巨噬细胞最初吞噬结核分枝杆菌会激活由种系编码的模式识别受体 (PRR) 组成的胞浆监视途径,导致 I 型干扰素 (IFN) 和促炎细胞因子产生增加、炎症小体活化和自噬 (2–4)。我们实验室和其他实验室的研究表明,结核分枝杆菌 DNA 和分枝杆菌衍生的环状二核苷酸可激活胞浆 DNA 传感途径 (5–8),从而驱动 I 型 IFN 的表达。虽然已经广泛研究了细胞浆病毒 RNA 在先天免疫感应中的作用,但细菌 RNA 对疾病发病机制的贡献尚不明确 (9)。最典型的 RIG-I 样受体 (RLR) 家族成员 RIG-I 和黑色素瘤分化因子 5 (MDA5) 包含一个中央 ATPase 含 DExD/H-box 解旋酶结构域和一个 C 末端阻遏物结构域,这两个结构域均参与 RNA 结合 (10, 11)。通过 RNA 结合激活后,2 个串联 caspase 激活和募集结构域 (CARD) 与衔接子线粒体抗病毒信号蛋白 (MAVS) 相互作用,介导 NF- κ B 和 IFN 调节因子 (IRF) 的诱导以及随后 IFN 刺激基因 (ISG) 的表达 (12–14)。尽管结构相似,RLR 检测的 RNA 种类往往不同,这些 RNA 种类往往具有病原体特异性,但不一定相互排斥 (11, 15)。越来越多的证据表明,RIG-I 在结核分枝杆菌感染的 I 型干扰素反应中起着非冗余作用 (16–18),它通过与特定的结核分枝杆菌 RNA 转录本结合,这些转录本利用分枝杆菌 ESX-1 分泌系统进入巨噬细胞胞质 (16)。我们最近发现,结核分枝杆菌 RNA 转录本能够通过与特定结核分枝杆菌 RNA 转录本结合,从而进入巨噬细胞胞质。

PRRSV通过双自噬受体P62和CCT2降低MDA5,以逃避抗病毒先天免疫
猪繁殖和呼吸综合征病毒(PRRSV)是一种主要的经济性病原体,已经发展了各种逃避先天免疫力的策略。抗病毒干扰素的下调在很大程度上通过利用细胞质黑色素瘤分化相关基因5(MDA5)来促进PRRSV免疫抗性,这是一种感受病毒RNA的受体。在这项研究中,观察到PRRSV感染中猪MDA5的下调转录和表达水平,并探索了详细的机制。我们发现,由于两个因素,p62和MDA5之间的相互作用得到了增强:上调的激酶CK2α和K63泛素化磷酸化受体p62的磷酸化修饰和由e3 Ubiquitinase Trim21催化的猪MDA5催化的猪MDA5的K63泛素化。由于这些修改,触发了经典的p62介导的自噬。此外,猪MDA5与含有TCP1亚基2(CCT2)的伴侣蛋白相互作用,该伴侣通过PRRSV NSP3增强。这种相互作用促进了独立于泛素化的MDA5-CCT2-NSP3的骨料形成和自噬清除率。总而言之,通过两种自噬途径在PRRSV感染中发生了增强的MDA5降解:MDA5与自噬受体p62和凝集受体CCT2的结合,导致强烈的先天免疫抑制。这项研究揭示了PRRSV感染中免疫逃避的一种新型机制,并为开发新疫苗或治疗策略提供了基本见解。

中链脂肪酸通过免疫调节受体GPR84
1型糖尿病(T1D)是一种自身免疫性疾病,导致胰腺β细胞破坏。coxsackievivirus B3(CVB3)感染和黑色素瘤分化相关蛋白5依赖性(依赖MDA5)抗病毒反应与T1D发育有关。IFIH1中的突变(编码为MDA5)与T1D易感性相关,但是这些突变如何促进T1D尚不清楚。Utilizing nonobese diabetic (NOD) mice lacking Ifih1 expression ( KO ) or containing an in-frame deletion within the ATPase site of the helicase 1 domain of MDA5 (Δ Hel1 ), we tested the hypothesis that partial or complete loss-of-function mutations in MDA5 would delay T1D by impairing proinflammatory pancreatic macrophage and T cell responses.在雌性点头和KO小鼠中开发的自发T1D类似,但在δHEL1小鼠中显着延迟,这可能部分是由于髓样衍生的抑制细胞同时增加。有趣的是,与点头小鼠相比,KO雄性小鼠自发性T1D增加了。虽然点头和KO小鼠产生了CVB3加速的T1D,而δHEL1小鼠则部分是由于I型IFN,胰腺浸润TNF +巨噬细胞,IFN-γ + CD4 + T细胞和perforin + CD8 + T细胞的部分保护。 此外,与野生型MDA5相比,δHEL1 MDA5蛋白减少了ATP水解。 我们的结果表明,MDA5功能受阻会延迟T1D,但MDA5的损失促进了T1D。虽然点头和KO小鼠产生了CVB3加速的T1D,而δHEL1小鼠则部分是由于I型IFN,胰腺浸润TNF +巨噬细胞,IFN-γ + CD4 + T细胞和perforin + CD8 + T细胞的部分保护。此外,与野生型MDA5相比,δHEL1 MDA5蛋白减少了ATP水解。我们的结果表明,MDA5功能受阻会延迟T1D,但MDA5的损失促进了T1D。

MDA5依赖性反应有助于自身免疫性糖尿病进展和阻碍
1型糖尿病(T1D)是一种自身免疫性疾病,导致胰腺β细胞破坏。coxsackievivirus B3(CVB3)感染和黑色素瘤分化相关蛋白5依赖性(依赖MDA5)抗病毒反应与T1D发育有关。IFIH1中的突变(编码为MDA5)与T1D易感性相关,但是这些突变如何促进T1D尚不清楚。Utilizing nonobese diabetic (NOD) mice lacking Ifih1 expression ( KO ) or containing an in-frame deletion within the ATPase site of the helicase 1 domain of MDA5 (Δ Hel1 ), we tested the hypothesis that partial or complete loss-of-function mutations in MDA5 would delay T1D by impairing proinflammatory pancreatic macrophage and T cell responses.在雌性点头和KO小鼠中开发的自发T1D类似,但在δHEL1小鼠中显着延迟,这可能部分是由于髓样衍生的抑制细胞同时增加。有趣的是,与点头小鼠相比,KO雄性小鼠自发性T1D增加了。虽然点头和KO小鼠产生了CVB3加速的T1D,而δHEL1小鼠则部分是由于I型IFN,胰腺浸润TNF +巨噬细胞,IFN-γ + CD4 + T细胞和perforin + CD8 + T细胞的部分保护。 此外,与野生型MDA5相比,δHEL1 MDA5蛋白减少了ATP水解。 我们的结果表明,MDA5功能受阻会延迟T1D,但MDA5的损失促进了T1D。虽然点头和KO小鼠产生了CVB3加速的T1D,而δHEL1小鼠则部分是由于I型IFN,胰腺浸润TNF +巨噬细胞,IFN-γ + CD4 + T细胞和perforin + CD8 + T细胞的部分保护。此外,与野生型MDA5相比,δHEL1 MDA5蛋白减少了ATP水解。我们的结果表明,MDA5功能受阻会延迟T1D,但MDA5的损失促进了T1D。
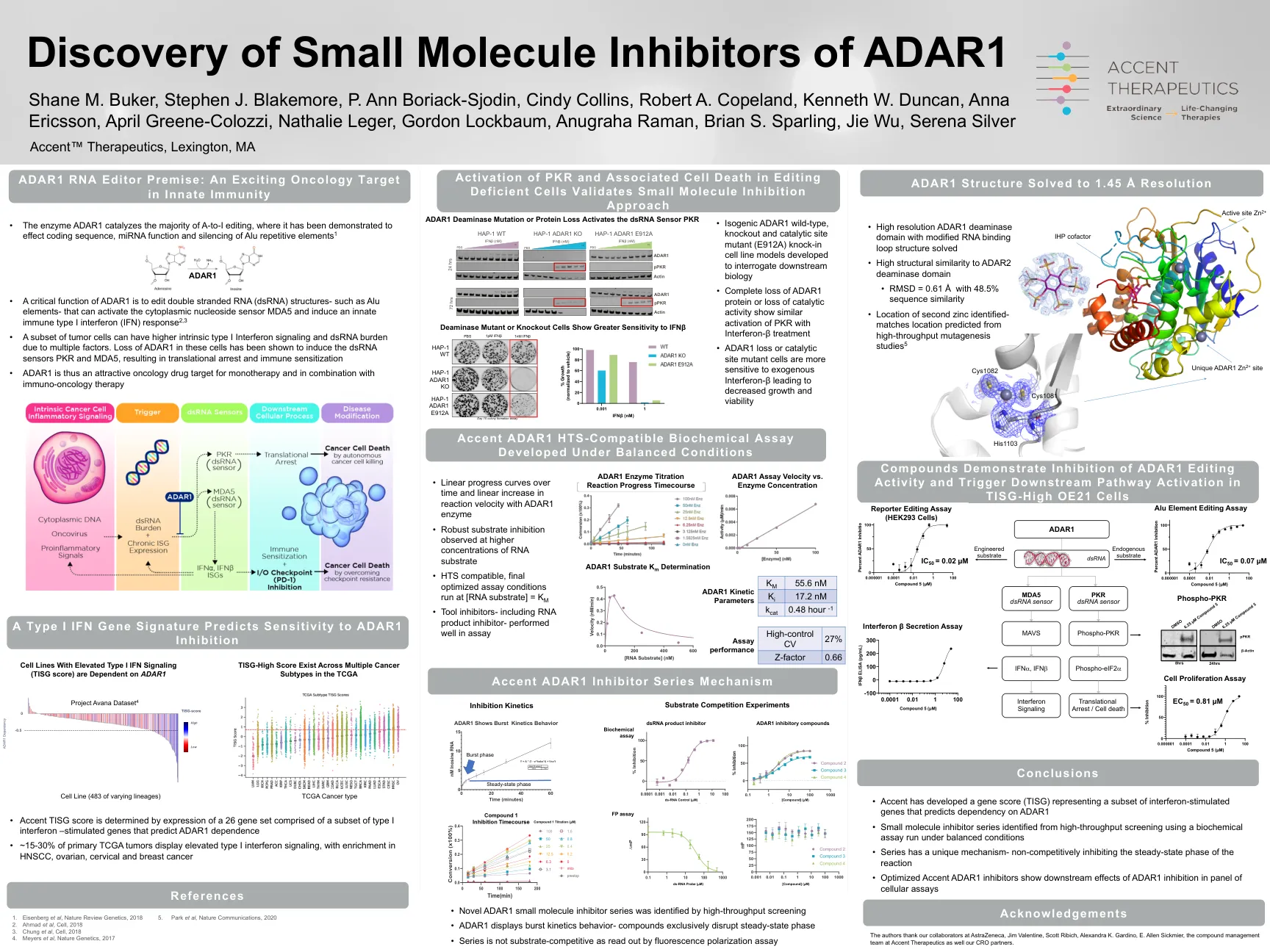

MDA5依赖性反应有助于自身免疫性糖尿病进展和阻碍lenvatinib或抗VEGF与抗PD-1结合使用...
引言黑色素瘤与所有其他恶性肿瘤相比,在晚期疾病中形成脑转移(BRM)的倾向最高(1)。免疫检查点抑制剂(ICI)已成为晚期黑色素瘤的中央治疗方法,并且仅在过去5年内进行了II期临床试验,才表明这些药物的颅内功效(2)。结合ipilimumab和nivolumab的最有效的ICI方案的响应率接近58%,但59%的个体发展了3-4级严重的不良事件(3)。对于那些最初没有反应,治疗进展或发展限制治疗的毒性(例如水肿或辐射坏死)的人,迫切需要替代方法。那些活跃的,以前未经治疗的BRM的人具有进一步的治疗局限性,因为许多试验仅允许那些患有小的,异常的转移或以前受辐照病变的试验。对穿透血脑屏障(BBB)并改善ICI活动的替代策略有直接且未满足的需求,而不会加剧毒性。最近的证据表明双重ICI和血管生成靶向的协同作用(4,5)。肿瘤利用增强的VEGF信号传导来逃避免疫系统并通过增加血管生成来促进生长(6)。用高剂量IL-2或ipilimumab治疗的黑色素瘤患者的生存率降低与较高的VEGF水平有关,因此,这表明可溶性VEGF可以用作临床结果的生物标志物,并具有逆转免疫抑制性肿瘤微型微环境(TME)的潜在靶标(TME)(7,8)。VEGF促进了Treg和髓样衍生的抑制细胞(MDSC)膨胀,抑制DC成熟,减轻效果T细胞反应,并改变淋巴细胞的运输到肿瘤中(9-14)。相反,血管生成抑制剂会促进T细胞浸润并减少免疫抑制肿瘤相关的巨噬细胞(TAMS)(15,16)。

通过RNA识别的先天免疫反应 - 生物化学
Tollike受体:对先天免疫的最新见解和观点。免疫,57,649 - 673。4)Rehwinkel,J。&Gack,M.U。(2020)RIG-I样受体:它们在RNA传感中的调节和作用。nat。修订版免疫。,20,537 - 551。5)Venkataraman,T.,Valdes,M.,Elsby,R.,Kakuta,S.,Cace- Res,G.,Saijo,S.,Iwakura,Y。,&Barber,G.N。(2007)DEXD/H盒RNA解旋酶LGP2的损失表现出不同的抗病毒反应。J. Immunol。 ,178,6444 - 6455。 6)Satoh,T.,Kato,H.,Kumagai,Y.,Yoneyama,M.,Sato,S.,Matsushita,K.,Tsujimura,T.,Fujita,T. (2010)LGP2是RIG-II和MDA5介导的抗病毒反应的积极调节剂。 proc。 natl。 学院。 SCI。 美国,107,1512 - 1517。 7)Bruns,A.M. (2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。 mol。 单元格,55,771 - 781。 8)乌鸦,Y.J. &Stetson,D.B。 (2022)I型干扰素:10年了。 nat。 修订版 免疫。 ,22,471 - 483。 9)村上,S。 (2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。 单元格,82,2236 - 2251。 10)Ablasser,A。 &Chen,Z.J。 (2019)CGA在行动中:在免疫和炎症中扩大角色。 Science,363,EAAT8657。J. Immunol。,178,6444 - 6455。6)Satoh,T.,Kato,H.,Kumagai,Y.,Yoneyama,M.,Sato,S.,Matsushita,K.,Tsujimura,T.,Fujita,T.(2010)LGP2是RIG-II和MDA5介导的抗病毒反应的积极调节剂。proc。natl。学院。SCI。 美国,107,1512 - 1517。 7)Bruns,A.M. (2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。 mol。 单元格,55,771 - 781。 8)乌鸦,Y.J. &Stetson,D.B。 (2022)I型干扰素:10年了。 nat。 修订版 免疫。 ,22,471 - 483。 9)村上,S。 (2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。 单元格,82,2236 - 2251。 10)Ablasser,A。 &Chen,Z.J。 (2019)CGA在行动中:在免疫和炎症中扩大角色。 Science,363,EAAT8657。SCI。美国,107,1512 - 1517。7)Bruns,A.M.(2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。mol。单元格,55,771 - 781。8)乌鸦,Y.J.&Stetson,D.B。(2022)I型干扰素:10年了。nat。修订版免疫。,22,471 - 483。9)村上,S。(2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。单元格,82,2236 - 2251。10)Ablasser,A。&Chen,Z.J。(2019)CGA在行动中:在免疫和炎症中扩大角色。Science,363,EAAT8657。11)Ablasser,A。&Hur,S。(2020)调节CGAS和RLR介导的对核酸的免疫力。nat。免疫。,21,17 - 29。12)Hopfner,K.P。&Hornung,V。(2020)CGAS刺信信号传导的分子机制和细胞功能。nat。修订版mol。细胞生物。 ,21,501 - 521。 13)伦纳德(J.N.),吉兰多(R. (2008)TLR3通过合作受体二聚体信号形式。 proc。 natl。 学院。 SCI。 美国,105,258 - 263。 14) (2008)带有双链RNA的Toll样重复3信号传导的结构基础。 Science,320,379 - 381。 15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。 (2005)Toll样受体3配体结合结构域的分子结构。 proc。 natl。 学院。 SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。细胞生物。,21,501 - 521。13)伦纳德(J.N.),吉兰多(R.(2008)TLR3通过合作受体二聚体信号形式。proc。natl。学院。SCI。 美国,105,258 - 263。 14) (2008)带有双链RNA的Toll样重复3信号传导的结构基础。 Science,320,379 - 381。 15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。 (2005)Toll样受体3配体结合结构域的分子结构。 proc。 natl。 学院。 SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。SCI。美国,105,258 - 263。14)(2008)带有双链RNA的Toll样重复3信号传导的结构基础。Science,320,379 - 381。15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。(2005)Toll样受体3配体结合结构域的分子结构。proc。natl。学院。SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。SCI。美国,102,10976 - 10980。16)Choe,J.,Kelker,M.S。和Wilson,I.A。(2005)人Toll样受体3(TLR3)外生域的晶体结构。科学,309,581 - 585。17)塔布塔(K.(2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。proc。SCI。SCI。natl。学院。美国,101,3516 - 3521。18)Davey,G.M.,Wojtasiak,M.,Proietto,A.I.,Carbone,F.R。,Heath,W.R。,&Bedoui,S。(2010)剪切边缘:CD8 T细胞免疫的启动:Surpes Simperx Simplex Virus 1型需要Cognate Tlr3在Vivo中的表达。J. Immunol。 ,184,2243 - 2246。 19)Oshiumi,H.,Okamoto,M.,Fujii,K.,Kawanishi,T.,Matsu-Moto,M.,Koike,S。,&Seya,T。(2011)TLR3/TICAM-1途径是对Poliovi-Rus Rus Infection的先天免疫反应的强制性。 J. Immunol。 ,187,5320 - 5327。 20)张,S.Y. (2007)疱疹患者的TLR3缺乏效率 -J. Immunol。,184,2243 - 2246。19)Oshiumi,H.,Okamoto,M.,Fujii,K.,Kawanishi,T.,Matsu-Moto,M.,Koike,S。,&Seya,T。(2011)TLR3/TICAM-1途径是对Poliovi-Rus Rus Infection的先天免疫反应的强制性。J. Immunol。 ,187,5320 - 5327。 20)张,S.Y. (2007)疱疹患者的TLR3缺乏效率 -J. Immunol。,187,5320 - 5327。20)张,S.Y.(2007)疱疹患者的TLR3缺乏效率 -

病例报告:探索托法替尼在调节...中的功效
一份病例报告强调了治疗一名 66 岁中国女性所面临的挑战,该女性被诊断为抗 MDA5 抗体阳性皮肌炎 (MDA5-DM),并伴有快速进展性间质性肺病 (RP-ILD)。尽管采取了积极的治疗干预措施,她的病情仍迅速恶化,凸显了这种亚型 DM 的严重性和破坏性。她的临床表现的一个显著特征是干扰素 (IFN)-g 和 IFN-a 水平显著升高,强调了 IFN 在驱动 MDA5-DM 相关 RP-ILD 的发病机制和进展中发挥的关键作用。为了阻止病情的持续进展,将 Janus 激酶 (JAK) 抑制剂托法替尼纳入她的治疗方案。这种治疗干预导致 IFN 相关细胞因子暂时减少,这为 JAK 抑制可以调节与该疾病有关的过度 IFN 反应带来了一线希望。其他四例类似病例强调了对 MDA5-DM 患者进行早期积极干预的重要性,以及使用 JAK 抑制剂阻断 IFN 的潜在治疗途径。迫切需要进行精心设计的临床试验,以揭示 RP-ILD 与 MDA5-DM 中 IFN 特征之间的复杂相互作用,并评估有望实现长期疗效和安全性的新型治疗靶点。