XiaoMi-AI文件搜索系统
World File Search SystemNakajima

湿件中的神经形态工程:最新进展及其前景
联合国大会(2015 年)制定了一项议程,其中包含 17 个目标,需要在全球范围内到 2030 年实现,以促进可持续的未来。实现这些目标需要设计和实施更有效的战略来管理复杂系统,包括人类及其社会、世界经济、城市地区、自然生态系统和气候(Gentili,2021a)。一项有前途的战略,即正在蓬勃发展的战略,依赖于人工智能 (AI) 和机器人技术的发展。人工智能帮助人类收集、存储和处理监测复杂系统不断演变所需的大数据(Corea,2019 年)。人工智能还帮助我们下定决心控制复杂系统的行为。硬机器人和软机器人让人类能够进入原本无法进入的环境。例如,它们帮助我们(1)研究其他行星的地球化学特征、考察海洋深渊以发现新的贵重材料和能源矿藏;(2)进入人体内部器官进行侵入性较小的手术;(3)在肮脏或危险的地方工作。开发人工智能的主要传统方法有两种(Lehman 等人,2014 年;Mitchell,2019 年)。第一种方法是编写在基于冯·诺依曼架构的电子计算机上运行的“智能”软件,该架构的主要缺点是处理单元和存储单元在物理上是分开的。一些软件模仿严谨的逻辑思维,而另一些软件模仿神经网络的结构和功能特征来学习如何从数据中执行任务。开发人工智能的第二种方法是在神经假体的硬件中实现人工神经网络,或设计类似大脑的计算机,将处理器和内存限制在同一空间中(所谓的内存计算;Sebastian 等人,2020 年)。如果人工神经网络由硅基电路或无机忆阻器制成,则它们是刚性的;如果基于有机半导体薄膜,则它们是柔性的(Christensen 等人,2022 年;Lee and Lee,2019 年;Wang 等人,2020 年;Zhu 等人,2020 年)。它们可以采用三种不同的架构进行设计:(A1)前馈(具有可训练的单向连接)、(A2)循环(具有可训练的反馈动作)或(A3)储层(由未训练的非线性动态系统与可训练的输入和输出层耦合而成)网络(Nakajima,2020 年;Tanaka 等人,2019 年;Cucchi 等人,2022 年;见图 1A)。在过去十年左右的时间里,一种开发人工智能的新颖而有前途的策略被提出:它包括通过湿件(即液体)中的分子、超分子和系统化学来模仿人类智能和所有其他生物所表现出的智能形式

20210604073646767_ja.pdf - 阿特拉斯有限公司
[1D2-OS-3a] 13:20-15:00 新闻媒体的数据科学 (1/3) (主席: Atsumu Sonoda) 1D2-OS-3a-01 线性和按需新闻视频分发分析广播中的使用风格(Masanori Takano、Yuki Ogawa、Fumiaki Taka、Soichiro Morishita)1D2-OS-3a-02 Twitter 上的政治分歧:以 2019 年参议院选举为例(Tsubasa Shindo、Yuki Ogawa、Yutaka Hattori)1D2 -OS-3a-03 使用基于 MMR 的句子选择和基于 TF-IDF 的句子压缩进行新闻文章摘要 (*Shotaro Ishihara、Norihiko Sawa) 1D2-OS-3a-04 利用 SNS 评论分析新闻媒体偏见(Shohei Hisada、Taichi Murayama、Juntaro Yada、Shoko Wakamiya、Eiji Aramaki)1D2-OS-3a-05 从汽车行业和社会的崩溃理解人工智能( Ryosuke Ozawa, Takeo Kiribayasi ) [1D3-OS-3b] 15:20-17:00 新闻媒体中的数据科学 (2/3) (主席: Yuki Ogawa ) 1D3-OS-3b- 01 (OS 特邀讲座) 新闻媒体中的问题日本政治交流:社会科学与数据科学的合作(Tetsuro Kobayashi)1D3-OS-3b-02 从推文中发现有争议的新闻文章的方法(Yui Fujikane、Kazuhiro Kazama、 Mitsuo Yoshida、Yoshinori Hijikata) 1D3-OS-3b-03 新闻服务中以内容多样性和标题为重点的用户参与度分析 (Atsumu Sonoda、Hiroto Nakajima、Fujio Toriumi) 1D3-OS-3b-04 量化新闻服务期间的消费者心理和行为使用文本挖掘研究 COVID-19 疫情/J-LIWC、J-MFD 和词共现网络的应用 (Kazutoshi Sasahara、Shinpei Okuda、Yu Igarashi) [1D4-OS-3c] 17:20-18:20 数据科学新闻媒体 (3/3) (主席:Masanori Takano) 1D4-OS-3c-01 根据用户关注者构成验证帖子传播情况 (Shogo Matsuno、Santi Seiyo、Takeshi Sakaki) , Yasuhiro Hino) 1D4-OS-3c-02 使用 BertSum 对日语新闻文章进行摘要总结的研究( Hideto Ishihara、Shotaro Ishihara、Hono Shirai)1D4-OS-3c-03 Twitter 上的新闻 浏览推文和观看视频之间的关系( Yuki Ogawa、Masanori Takano、Soichiro Morishita、Fumiaki Taka)### 会场 E OS 会场 3 ### [1E2-OS-2] 13:20-14:40 认知偏差・多重解释和人工智能(1/1)(主席:Shohei Hidaka) 1E2-OS-2-01 麻木错觉与自我触摸错觉之间的权衡研究 (Yutaro Sato、Godai Saito)、Kotaka Kenri 1E2-OS-2-02 为什么狼人杀中人会被愚弄?/从认知偏差的角度进行思考(Kanzen Noriaki、Takeshi Ito)1E2-OS-2-03 从对新事物的态度看信念偏差的出现冠状病毒机制(Daiki Kondo)
![利用人工智能(AI)技术与电磁学进行优化设计 [I]](/simg/e\e2b202296b0567a88b2efb733a168494aa2a8937.png)
利用人工智能(AI)技术与电磁学进行优化设计 [I]
(1) MP Bendsøe 和 N. Kikuchi,“使用均质化方法在结构设计中生成最佳拓扑”,Comp. Methods in Appl. Mech. Eng.,第 71 卷,第 197-224 页,1988 年。 (2) MP Bendsøe 和 O. Sigmund,拓扑优化,理论、方法和应用,Springer,2004 年。 (3) Hidenori Sasaki 和 Hajime Igarashi,“使用傅里叶级数对 IPM 电机进行拓扑优化”,Journal of Electrical Engineering (B),第 137 卷,第 3 期,第 245-253 页,2017 年 3 月。 (4) Y. Tsuji 和 K. Hirayama,“使用基于函数扩展的折射率分布的拓扑优化方法设计光路设备”,IEEE Photonics Technol. Lett., (5) T. Sato、H. Igarashi、S. Takahashi、S. Uchiyama、K. Matsuo 和 D. Matsuhashi,“使用拓扑优化实现内置永磁同步电机转子形状优化”,《电气工程杂志 (D)》,第 135 卷,第 3 期,第 291-298 页,2015 年 3 月。 (6) S. Kobayashi,“实数编码 GA 的前沿”,《人工智能杂志》,第 24 卷,第 1 期,第 147-162 页,2009 年 1 月。 (7) T. Sato、K. Watanabe 和 H. Igarashi,“基于正则化高斯网络的电机多材料拓扑优化”,《IEEE 会刊》, (8) S. Hiruma、M. Ohtani、S. Soma、Y. Kubota 和 H. Igarashi,“参数和拓扑优化的新型混合:应用于永磁电机,”IEEE Trans. Magn.,第 57 卷,第 7 期,8204604,2021 年 (9) Y. Otomo 和 H. Igarashi,“用于无线电源传输设备的磁芯 3-D 拓扑优化,”IEEE Trans. Magn.,第 55 卷,第 6 期,8103005,2019 年。 (10) K. Itoh、H. Nakajima、H. Matsuda、M. Tanaka 和 H. Igarashi,“使用带归一化高斯网络的拓扑优化开发用于缝隙天线的小型介电透镜,”IEICE Trans. Electron., E101-C 卷,第 10 期,第 784-790 页,2018 年 10 月。 (11) N. Hansen、SD Müller 和 P. Koumoutsakos,“通过协方差矩阵自适应降低去随机化进化策略的时间复杂度(CMA-ES),”进化计算,第 11 卷,第 1 期,第 1-18 页,2003 年。 (12) N. Aage、E. Andreassen、BS Lazarov 和 O. Sigmund,“用于结构设计的千兆体素计算形态发生”,自然,第 550 卷,23911,2017 年。

引用Liu L,Tang Y,Shao J,Fan B,Yang Y,Zhang Y,Zhao X,Xue H,Sun H,Sun H,Zhang X,Zhang X,Zhang Y和Xu B(2025)培养文化中阶段依赖的变化
绵羊。 这种差异对尖端生殖生物技术的应用具有深远的影响,并可能阻碍高质量母猪生殖性能的改善和建立人类疾病的猪模型。 因此,猪卵母细胞IVM的优化已成为全球猪繁殖群落研究的关键领域。 除了激素水平(Lu等,2014; Sakaguchi和Nagano,2020),氨基酸的可用性(Bahrami等,2019; Lee等,2019),以及抗氧化剂补充剂(Das等,2014; li等,2019; li et al。卵母细胞成熟质量的重要决定因素(Baltz和Zhou,2012年)。 超过一个世纪的哺乳动物胚胎培养经验强调了细胞体积控制在确定植入前胚胎的发育轨迹中的关键作用(Biggers,1998)。 早期培养哺乳动物胚胎的努力是基于仿生型的,在培养基中定位了受精卵的卵子,其渗透压近似于该生物体内部环境(290 - 310 MOSM)。 然而,这种方法导致物种特定的胚胎停滞,归因于渗透条件(Goddard和Pratt,1983; Camous等,1984; Camous等,1984; Bolton等,1989; Kishi等,1991)。 值得注意的是,成功克服了这种发育障碍的培养基要么将培养基的渗透压降低,要么融合了有机渗透剂,例如甘氨酸(Gly),Betaine,β-丙氨酸和谷氨酰胺,渗透性为310 MOSM的培养基(Van Winkle等,1990; Biggers et al eal and osmolartials osmolarity。绵羊。这种差异对尖端生殖生物技术的应用具有深远的影响,并可能阻碍高质量母猪生殖性能的改善和建立人类疾病的猪模型。因此,猪卵母细胞IVM的优化已成为全球猪繁殖群落研究的关键领域。除了激素水平(Lu等,2014; Sakaguchi和Nagano,2020),氨基酸的可用性(Bahrami等,2019; Lee等,2019),以及抗氧化剂补充剂(Das等,2014; li等,2019; li et al。卵母细胞成熟质量的重要决定因素(Baltz和Zhou,2012年)。超过一个世纪的哺乳动物胚胎培养经验强调了细胞体积控制在确定植入前胚胎的发育轨迹中的关键作用(Biggers,1998)。早期培养哺乳动物胚胎的努力是基于仿生型的,在培养基中定位了受精卵的卵子,其渗透压近似于该生物体内部环境(290 - 310 MOSM)。然而,这种方法导致物种特定的胚胎停滞,归因于渗透条件(Goddard和Pratt,1983; Camous等,1984; Camous等,1984; Bolton等,1989; Kishi等,1991)。值得注意的是,成功克服了这种发育障碍的培养基要么将培养基的渗透压降低,要么融合了有机渗透剂,例如甘氨酸(Gly),Betaine,β-丙氨酸和谷氨酰胺,渗透性为310 MOSM的培养基(Van Winkle等,1990; Biggers et al eal and osmolartials osmolarity。例如,已证明在KSOM或CZB培养基中培养小鼠胚胎(250 - 275 MOSM)可以抵御两细胞停滞(Chatot等,1990; Lawitts and Biggers,1991; 1993; 1993; Hadi等,2005)。当受外部条件干扰时,细胞体积控制的迅速恢复是通过Na + /H +交换器NHE1和HCO 3 + /Cl- -Chressanger AE2的激活来介导的,该E2调节Na +和Cl-的细胞内浓度。尽管如此,至关重要的是避免过度高离子浓度,这可能破坏正常的细胞生理和生化过程。Subsequently, preimplantation embryos and oocytes reactivate speci fi c organic osmolyte channels to internalize uncharged osmolytes, replacing inorganic ions and ensuring that cells maintain normal physiological and biochemical processes ( Alper, 2009 ; Donowitz et al., 2013 ; Nakajima et al., 2013 ; Tscherner et al., 2021)。对小鼠卵母细胞中的细胞体积调节机制的研究表明,编码Gly Transporter的SLC6A9的特定缺失消除了植入前胚胎中的GLY转运及其对催眠应激的能力(Tscherner等人,2023)。这些发现强调了对哺乳动物卵母细胞和植入前胚胎的健康发展进行精确细胞体积调节的必要性。gly是蛋白质和核酸合成中必不可少的前体,这对于快速细胞增殖至关重要(Redel等,2016; Alves等,2019)。据报道,Gly是猪卵泡液中最丰富的氨基酸(Hong and Lee,2007),这表明Gly可能是在体外改善卵母细胞成熟的重要因素。虽然精确的机制仍有待完全阐明,但新出现的证据表明,Gly作为牛胚胎和小鼠卵母细胞发展中的有机渗透剂的重要作用(Zhou等,2013; Herrick et al。
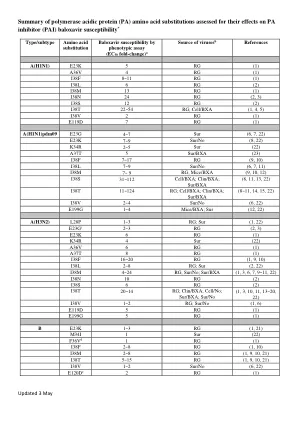
摘要 - 聚合酶 - 酸性 - (PA) - 蛋白质 - 氨基 -
* PA中的其他氨基酸取代,在参考文献1(Omoto S等,2018)和#2(Hashimoto T等,2020年)中研究了Baloxavir易感性没有变化的其他氨基酸取代。通过基于细胞培养的测定法评估(焦点,斑块或屈服分析,高含量成像中和(提示)和ViroDot分析)。EC 50倍变化。b细胞,细胞培养;临床试验;小鼠,鼠标模型; RG,反向遗传学; SUR,监视研究; BXA,在Baloxavir压力下选出的取代;不,Baloxavir不使用。c e23g(T0831)。通过表型测定测试了带有E23G的RG病毒。d对应于A36V A型A型PA中的A36V。 E对应于A型A型PA中的E119D。参考文献1。Omoto S,Speranzini V,Hashimoto T,Noshi T,Yamaguchi H,Kawai M,Kawaguchi K,Uehara T,Shishido T,Naito A,Naito A,Cusack S.2018。通过核酸内切酶抑制剂Baloxavir maroxil诱导的流感病毒变体的表征。SCI REP 8:9633。2。Hashimoto T,Baba K,Inoue K,Okane M,Hata S,Shishido T,Naito A,Wildum S,Omoto S.2020。在Baloxavir Marboxil的临床试验中检测到的流感病毒的三聚体RNA聚合酶复合物中氨基酸取代的全面评估。流感其他呼吸病毒DOI:10.1111/irv.12821。3。ince WL,Smith FB,O'Rear JJ,Thomson M.2020。J Infect DIS 222:957-961。 4。 2018。J Infect DIS 222:957-961。4。2018。治疗 - 伴随流感病毒聚合酶酸性取代率与Balosavir Maroxavir Marboxil试验中的i38中的i38中的酸性取代相关。Noshi T, Kitano M, Taniguchi K, Yamamoto A, Omoto S, Baba K, Hashimoto T, Ishida K, Kushima Y, Hattori K, Kawai M, Yoshida R, Kobayashi M, Yoshinaga T, Sato A, Okamatsu M, Sakoda Y, Kida H, Shishido T, Naito A.Baloxavir酸的体外表征,Baloxavir酸是一种流感病毒聚合酶PA亚基的第一类帽依赖性内切酶抑制剂。抗病毒Res 160:109-117。5。Takashita E,Morita H,Ogawa R,Nakamura K,Fujisaki S,Shirakura M,Kuwahara T,Kishida N,Watanabe S,Odagiri T.2018。流感病毒对新型帽依赖性核酸内切酶抑制剂baloxavir maroxil的敏感性。前微生物9:3026。6。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。 2019。 评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。 欧元监视24:1800666。 7。 Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A. 2020。 全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。 抗病毒Res 175:104718。 8。 2020。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。2019。评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。欧元监视24:1800666。7。Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A.2020。全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。抗病毒Res 175:104718。8。2020。Takashita E, Abe T, Morita H, Nagata S, Fujisaki S, Miura H, Shirakura M, Kishida N, Nakamura K, Kuwahara T, Mitamura K, Ichikawa M, Yamazaki M, Watanabe S, Hasegawa H, Influenza Virus Surveillance Group of J.流感A(H1N1)PDM09病毒,由于未经Baloxavir治疗的儿童检测到PA E23K替代而表现出对Baloxavir的敏感性降低。抗病毒Res 180:104828。 9。 Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。 2019。 亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。 抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 180:104828。9。Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。2019。亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 164:91-96。10。Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。2020。流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。Proc Natl Acad Sci U S A 117:8593-8601。11。Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。2020。季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。J Infect DIS 221:367-371。12。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y.2020。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。J Infect Dis 221:1699-1702。13。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。J Infect DIS 222:121-125。14。J Infect DIS 221:63-70。15。2020。16。在2018-2019流感季节治疗流感A的儿童后,检测Baloxavir Marboxil易感性降低的变体。Checkmahomed L,M'Hamdi Z,Carbonneau J,Venable MC,Baz M,Abed Y,Boivin G.2020。抗性抗性聚合酶酸I38T取代对当代流感A(H1N1)PDM09和A(H3N2)菌株的适应性的影响。Imai M, Yamashita M, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kiso M, Murakami J, Yasuhara A, Takada K, Ito M, Nakajima N, Takahashi K, Lopes TJS, Dutta J, Khan Z, Kriti D, van Bakel H, Tokita A, Hagiwara H, Izumida N,Kuroki H,Nishino T,Wada N,Koga M,Adachi E,Jubishi D,木谷H,Kawaoka Y.流感A的变体降低了对日本患者分离的Baloxavir敏感性的变体,并通过呼吸道液滴进行拟合。NAT微生物5:27-33。 Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza VirusNAT微生物5:27-33。Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza Virus