XiaoMi-AI文件搜索系统
World File Search SystemNeurons

神经元信息整合同步的进化起源
脑表达基因的进化速度明显慢于其他组织中表达基因的进化速度,这一现象可能是由于高级功能限制造成的。其中一个限制可能是神经元组合对信息的整合,从而增强环境适应性。本研究通过三种类型的同步探索了神经元中信息整合的生理机制:化学、电磁和量子。化学同步涉及多巴胺和乙酰胆碱等神经递质的弥散释放,导致传输延迟数毫秒。电磁同步包括动作电位、电间隙连接和偶联。电间隙连接使皮质 GABA 能网络内的快速同步成为可能,而偶联则使轴突束等结构能够通过细胞外电磁场同步,速度超过了化学过程的速度。据推测,量子同步涉及离子通道通过期间的离子相干性和髓鞘内光子的纠缠。与化学和电磁过程中的有限时间同步不同,量子纠缠提供瞬时非局部相干状态。神经元可能从较慢的化学扩散进化为快速的时间同步,离子通过皮质 GABAergic 网络内的间隙连接可能促进快速伽马波段同步和量子相干。这篇小综述汇编了有关这三种同步类型的文献,为解决神经元组装中结合问题的生理机制提供了新的见解。

安全访问租户管理指南的神经元
END USER LICENSE AGREEMENT 7 Preface 8 Document conventions 8 Revision History 10 Ivanti Neurons for Secure Access Overview 11 What's New 14 Version 22.8R1 14 Version 22.7R1.6 14 Version 22.7R1.5 15 Version 22.7R1.4 16 Version 22.7R1.3 16 Version 22.7R1.2 17 Version 22.7R1 17 Version 22.6R1 18 Version 22.5R1 20 Version 22.4R3 20 Version 22.4R1 21 Logging in to Ivanti Neurons for Secure Access 22 Preparing to Log in 22 Logging in to the Ivanti Neurons for Secure Access as a Tenant Admin 22 Logging out of the Ivanti Neurons for Secure Access 28 Managing Existing Ivanti Connect Secure 9.x Appliance 29 Registering Ivanti Connect Secure Gateway 30 Completing Registration of an ICS Appliance 33 Reregistration of ICS Gateway 35 Ivanti Connect安全网关部署37部署在VMware上37在Hyper-V 38上部署在KVM上的部署在KVM上38部署在AWS Cloud上38部署在Azure Cloud 45上的部署在Azure Cloud 45上部署在Google Cloud Cloud Platform上48 IVANTI 48 IVANTI IVANTI ivanti Secuthway Anallytics 49 Recection 49 Recess Intervion 49综合活动53综合活动53活动103审查网关状态和版本109检查日志116配置可行的见解129生成报告132管理会话139
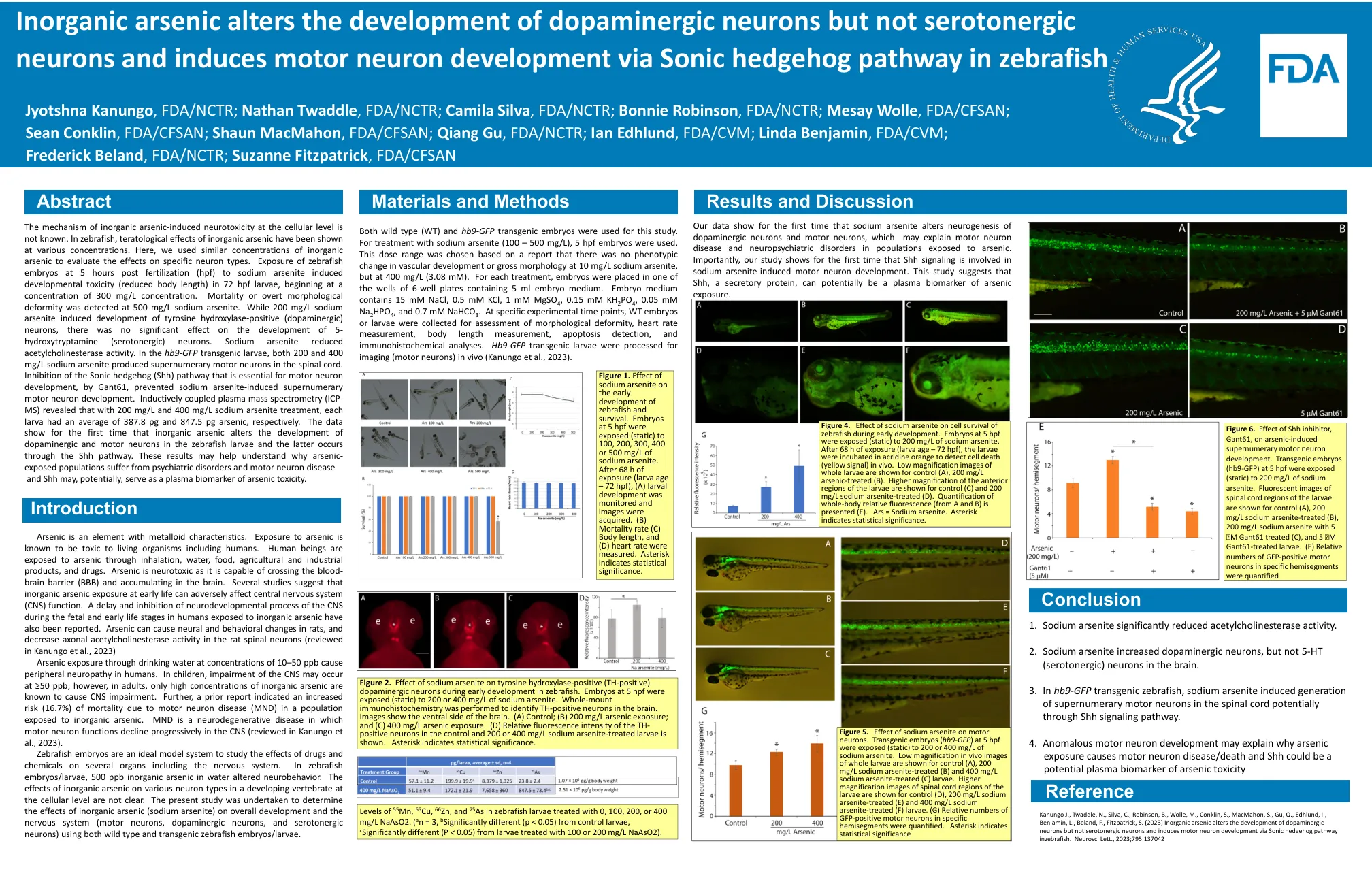
无机砷改变多巴胺能神经元的发育,但不改变血清素能神经元的发育,并通过 Sonic Hedgehog 通路诱导斑马鱼的运动神经元发育
无机砷在细胞水平上诱发神经毒性的机制尚不清楚。在斑马鱼中,不同浓度的无机砷均有致畸作用。在这里,我们使用了类似浓度的无机砷来评估其对特定神经元类型的影响。受精后 5 小时 (hpf) 的斑马鱼胚胎暴露于亚砷酸钠中,在 72 hpf 幼虫中诱发发育毒性(体长缩短),浓度从 300 mg/L 开始。在 500 mg/L 亚砷酸钠下检测到死亡或明显的形态畸形。虽然 200 mg/L 亚砷酸钠诱导酪氨酸羟化酶阳性(多巴胺能)神经元的发育,但对 5-羟色胺(血清素能)神经元的发育没有显著影响。亚砷酸钠降低了乙酰胆碱酯酶活性。在hb9-GFP转基因幼鱼中,200和400mg/L亚砷酸钠均在脊髓中产生了多余的运动神经元。通过Gant61抑制运动神经元发育所必需的Sonic Hedgehog(Shh)通路,可以阻止亚砷酸钠诱导的多余运动神经元发育。电感耦合等离子体质谱(ICP-MS)分析表明,在200mg/L和400mg/L亚砷酸钠处理下,每只幼鱼平均砷含量分别为387.8pg和847.5pg。数据首次表明无机砷改变斑马鱼幼鱼多巴胺能神经元和运动神经元的发育,后者是通过Shh通路发生的。这些结果可能有助于理解为什么接触砷的人群会患上精神疾病和运动神经元疾病,并且 Shh 可能潜在地充当砷毒性的血浆生物标志物。

丘脑神经元中突触基因表达和回路的共同保守性
。CC-BY-NC-ND 4.0 国际许可下可用(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者此版本于 2025 年 1 月 23 日发布。;https://doi.org/10.1101/2025.01.23.634521 doi:bioRxiv 预印本

人类皮质神经元的体外全细胞电生理学数据集
1 克雷姆比尔神经信息学中心,成瘾和精神健康中心,加拿大安大略省多伦多,M5T 1R8 2 克雷姆比尔脑研究所,大学健康网络,加拿大安大略省多伦多,M5T 1M8 3 多伦多大学特默蒂医学院医学科学研究所,加拿大安大略省多伦多,M5S 1A8 4 推进神经技术创新应用中心(CRANIA),加拿大安大略省多伦多,M5S 1A4 5 多伦多大学外科系,神经外科分部,加拿大安大略省多伦多,M5T 1P5 6 多伦多大学生物医学工程研究所,加拿大安大略省多伦多,M5S 3G9 7 多伦多大学电气和计算机工程系,加拿大安大略省多伦多,M5S 3G8 8 马克斯普朗克 - 多伦多大学神经科学与技术中心,加拿大安大略省多伦多,M5S 1A4 9 中心促进神经技术创新应用(CRANIA)的加拿大多伦多大学精神病学系, ...

哺乳动物脑中多晶镜神经元的突触和电路功能
哺乳动物脑中的神经元不限于释放单个神经递质,而是通常将神经递质的神经递质释放到突触后细胞上。在这里,我们回顾了整个哺乳动物中枢神经系统中发现的多晶月神经元的最新发现。我们重点介绍了最新的技术创新,这些创新使新的多晶镜神经元及其突触特性的研究成为可能。我们还专注于轴突末端和突触囊泡上神经递质corelease所需的机制和分子成分,以及多种晶状体神经元在多种脑电路中的一些可能功能。我们期望这些方法将导致对多晶镜神经元的机制和功能的新见解,它们在电路中的作用以及它们对正常和病理大脑功能的贡献。

镜像神经元:最新发现和进一步的观点
反映神经元被认为是与他人建立联系的能力,而不是有意识的水平,通过模仿,理解和提供帮助来学习;共情。这些连接不是直接的,而是根据一个人的经验进行介导的[1]。镜像神经元在儿童时期很重要,它们对于学习和获取新技能非常重要。他们参与思考,计划,控制和记忆。如果孩子观察到动作,镜像神经元将激活并形成新的神经联系,就好像他或她亲自进行了动作一样。镜像神经元的有效功能可在任何领域,更大的情绪智力和更高的同理心[1]带来出色的发展。

零信任访问入门指南的神经元
End User Agreement 5 Preface 6 Document conventions 6 Text formatting conventions 6 Requesting Technical Support 8 Self-Help Online Tools and Resources 8 Opening a Case with Support 8 Reporting Documentation Issues 8 What's New 9 Version 22.6R1 9 Version 22.5R1 9 Version 22.4R3 10 Getting Started with Ivanti Neurons for Zero Trust Access 11 What is nZTA 11 Deploying and Using nZTA 11 Manually Configuring Your nZTA部署13创建用户身份验证服务15工作流程:创建本地身份验证策略16工作流程:与Azure AD AD AD 26工作流程创建SAML身份验证策略26工作流程:使用本地ICS创建SAML身份验证策略43工作流程43工作流程:将TOTP添加到身份验证策略63中的用户组73与用户组合78启用78的启用78 Inders Offect 78 Indust Inderion 78 Inders Offect of Demant 78 Submistion 84 Inders Exprion 74 Gateway in VMware vSphere 89 Workflow: Creating a Gateway in Amazon Web Services 98 Workflow: Creating a Gateway in Microsoft Azure 108 Workflow: Creating a Gateway in KVM/OpenStack 126 Workflow: Creating a Gateway in Google Cloud Platform 141 Workflow: Creating a Gateway in Oracle Cloud Platform 163 Next Steps 261 Creating Device Policies and Device Rules 262 Introduction 262 Creating Device Policies 264 Creating Device Rules 268 Next Steps 284创建应用程序和应用程序组285简介285将应用程序添加到控制器285将应用程序组添加到控制器289下一步291

运动神经元和娱乐方式如何支持身体发育
虽然运动技能从出生开始似乎是自然发展的,但实际上是婴儿的早期经历,才真正助长了这一发展。定期经历大量互动的婴儿(例如通过身体上的互动)将从这些互动中发展他们的运动技能。但是,对于那些没有经常互动或经历过幼儿创伤的孩子,其运动神经元建立长期联系的能力降低了,结果他们的发展也是如此。研究发现,儿童不良经历的儿童经历运动技能开发受损的可能性要高出五到七倍。
