XiaoMi-AI文件搜索系统
World File Search SystemNeurons

MDM的Ivanti神经元96管理员指南
About Ivanti Neurons for MDM 5 New features summary 6 General features and enhancements 6 Android features 7 iOS, macOS, and tvOS features 8 Windows features 11 Mobile Threat Defense features 11 Getting Started 12 Solution Overview 12 Setting preferred language in a browser 18 Unified navigation interface for Ivanti Neurons for MDM and Access 19 Device Admin (DA) mode to manage Android devices - deprecated 19 Configuring macOS devices 21 Configuring and using registration confirmation emails 26 Configuring and using policy compliance notification emails 27 On-demand features 29 Preparing for Android Enterprise device support 33 Dashboard 36 Working with Widgets 37 App Insights 52 Using Scheduled Reports 58 Using Custom Reports 69 Users 80 Adding Users 81 User Groups 87 User Settings 91 User Branding 107 User Enrollment with Apple Business Manager 109 Account driven User Enrollment 121 User Licenses 123 Managing用户124个设备168从设备开始使用169个设备组188非管理设备195 App库存197管理设备201 Apps 288

直接重新编程对诱导神经元的ins和uns
了解细胞类型的特定转录因子已促进了细胞重编程方法的进展,例如将体细胞直接重编程为诱导的神经元(IN)。直接重编程的方法需要神经元允许通过神经元特异性microRNAS确定基因激活,关键神经元信号通路的化学调节或通过病毒载体过表达的化学调节,并具有一些重编程策略,需要将这些方法组合来诱导神经元电池效果。这些方法已用于多种细胞类型,包括纤维细胞,肝细胞,外周血单核和T细胞。从皮肤活检和血液样本中创造的能力以及人工诱导的年龄和疾病相关的表型的最新进展正在加速迟到神经退行性疾病的疾病模型的发展。在这里,我们回顾了神经元转录组的激活如何改变供体细胞的表观遗传景观,以促进对神经元的重编程。我们还讨论了使用DNA结合结构域(例如CRISPR/DCAS9)通过激活内源性神经元细胞纳入确定基因来诱导神经元细胞命运的优势来诱导神经元细胞命运。

在发育中的Claustrum中,神经元的独特“反向”迁移
这篇早期版本的文章已经过同行评审和接受,但尚未通过构图和复制过程。最终版本的样式或格式可能会略有不同,并且将包含指向任何扩展数据的链接。

remelelination保护神经元免受DLK介导的神经变性
慢性脱髓鞘和少突胶质细胞丧失剥夺神经元的关键支持。正是神经元的变性及其连接导致脱髓鞘疾病的逐渐残疾。但是,慢性脱髓鞘是否触发神经变性及其如何触发。我们表征了两种诱导脱髓鞘的遗传小鼠模型,一种是有效的再髓样区别,另一个是通过雷诺德式衰竭和慢性脱髓鞘而区分的。虽然两种脱髓鞘线都具有轴突损伤,但具有阻塞的透明度的小鼠具有升高的神经性凋亡,并且微胶质细胞炎症的改变,而具有效率的透明性的小鼠没有神经元细胞凋亡,并且功能恢复改善。rem髓无能力的小鼠表明,双亮氨酸拉链激酶(DLK)下游激活增加了神经元核中C-JUN的磷酸化。DLK阻滞C-Jun磷酸化和脱髓鞘神经元的凋亡的药理抑制或遗传疾病。一起,我们证明了与神经保护作用相结合,并确定DLK抑制作用是慢性脱髓鞘神经元的保护策略。
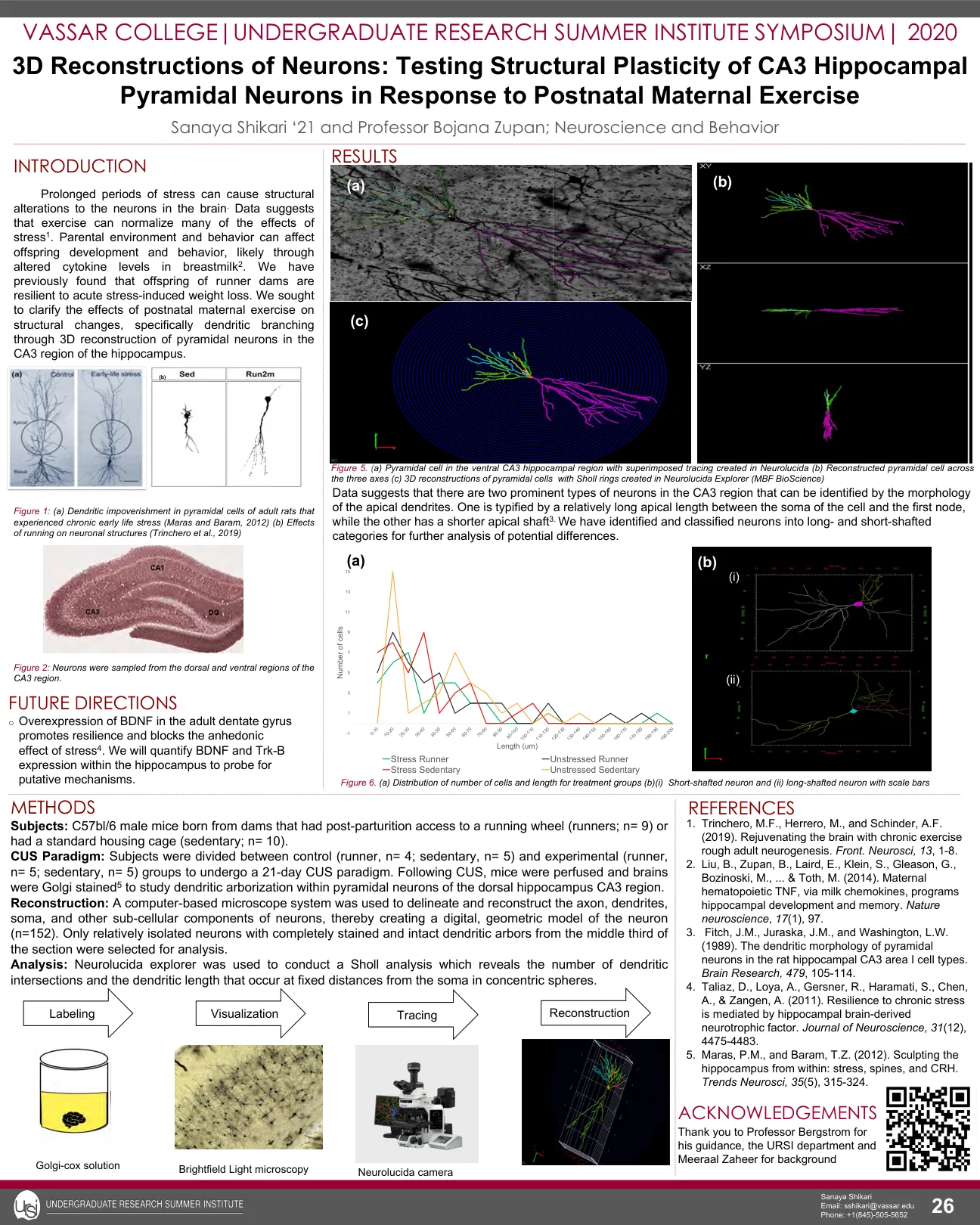
神经元的 3D 重建:测试 CA3 海马锥体神经元对产后母亲运动的反应的结构可塑性
方法 受试者:C57bl/6雄性小鼠,其母鼠产后可使用跑轮(跑步者;n= 9)或使用标准笼子(久坐;n= 10)。 CUS 范式:将受试者分为对照组(跑步者,n= 4;久坐组,n= 5)和实验组(跑步者,n= 5;久坐组,n= 5),接受为期 21 天的 CUS 范式。 CUS 之后,对小鼠进行灌注,并对大脑进行 Golgi 染色 5,以研究背海马 CA3 区锥体神经元内的树突树枝状化。 重建:使用基于计算机的显微镜系统来描绘和重建神经元的轴突、树突、胞体和其他亚细胞成分,从而创建神经元的数字几何模型(n=152)。仅选择切片中间三分之一处具有完全染色和完整树突状体的相对分离的神经元进行分析。分析:使用 Neurolucida explorer 进行 Sholl 分析,该分析揭示了同心球中距胞体固定距离处出现的树突交叉点数量和树突长度。

背部后顶神经元的时间细节
研究文章|行为/认知混合选择性编码内容 - 时间细节由背部后壁神经元https://doi.org/10.1523/jneurosci.1677-23.2023收到:2023年9月5日收到:2023年10月27日接受:2023年10月1日接受:2023年11月1日接受:

计数神经元:侵入性和非侵入性技术的比较
本文讨论了神经科学研究中用于计数神经元和估计其密度的各种技术(分为侵入性和非侵入性)。侵入性技术包括物理移除和染色脑组织以计数神经元,而非侵入性技术则可以检查活体动物或人类的大脑结构和功能。侵入性技术的例子包括立体学、光学分馏器、手动细胞计数、共聚焦显微镜和电子显微镜。非侵入性技术的例子包括磁共振成像 (MRI)、脑电图 (EEG) 和功能性近红外光谱 (fNIRS)。表 1 总结了每种技术的优缺点。重点介绍了用于估计神经元密度的非侵入性 NIR 光谱技术的最新进展。未来的研究和技术进步,尤其是在非侵入性技术方面的进步,可以让我们更好地了解神经回路、它们的功能以及我们的日常生活。

人类大脑中单个神经元的意志控制
脑机接口使神经科学家能够将特定的神经活动模式与特定的行为联系起来。因此,除了目前的临床应用外,脑机接口还可用作研究大脑学习和可塑性的神经机制的工具。数十年来使用此类脑机接口的研究表明,动物(非人类灵长类动物和啮齿动物)可以通过操作条件反射自我调节大脑各种运动相关结构的神经活动。在这里,我们要问的是,人类大脑是一个由超过 800 亿个神经元组成的复杂互连结构,它能否学会在最基本的层面——单个神经元——上自我控制。我们利用这个独特的机会记录了 11 名癫痫患者的单个单元,以探索边缘系统和其他与记忆相关的大脑结构中单个(直接)神经元的发放率是否可以受到意志控制。为此,我们开发了一个视觉神经反馈任务,训练参与者通过调节他们大脑中任意选择的神经元的活动来移动屏幕上的方块。值得注意的是,参与者能够有意识地调节这些以前未经研究的结构中的直接神经元的发放率。我们发现一部分参与者(学习者)能够在一次训练课程中提高他们的表现。成功的学习的特点是:(i)直接神经元的高度特异性调节(表现为发放率和爆发频率显著增加);(ii)直接神经元的活动与邻近神经元的活动同时去关联;(iii)直接神经元与局部 alpha/beta 频率振荡的稳健锁相,这可能为促进这种学习的潜在神经机制提供一些见解。记忆结构中神经元活动的意志控制可能为探索人类记忆的功能和可塑性提供新方法,而无需外部刺激。此外,这些大脑区域神经活动的自我调节可能为开发新型神经假体提供途径,用于治疗通常与这些大脑结构中的病理活动相关的神经系统疾病,例如药物难治性癫痫。

解开神经元群体之间的信号流
采用这种概念,一些跨区域研究比较了刺激呈现后跨区域神经反应开始的时间 13 – 15 或归因于自上而下过程的选择性出现的时间 16 – 20 。其他研究利用同步记录,通过成对脉冲相关性 21 – 26 和信息论测量 27 测量了两个区域之间的时间延迟。同样,局部场电位的跨区域相位延迟也被测量了 28 – 31 。这些基于时间的方法增进了我们对信号如何在大脑区域间传播的理解。然而,由于这些方法主要关注神经元对或神经活动的总体测量,因此关于神经元群体如何协调其活动以实现跨区域信号传导仍有许多未知之处。

小胶质细胞,星形胶质细胞和神经元中的细胞类型特异性激活
相应的作者Dietmar R.神经病理学的Thal实验室O&N IV Hestraat 49 -Bus 1032 3000 Leuven Tel。:0032/16/3-44047电子邮件地址:dietmar.thal@kuleuven.be