XiaoMi-AI文件搜索系统
World File Search SystemRAD51

全基因组的DNA修复蛋白分析鉴定单细胞中的高阶配位
a,实验设置和集成的概述。b,染色体1p上的信号。左:在 +DSB条件下的单细胞热图(RPKM),其顶部为 +DSB(有色)和–DSB(灰色)条件的单细胞聚集体。右:带有覆盖MSR调用的单细胞线图。asisi图案,用黑线注释,红色三角形表示经常裂解(或“顶部”)位点。c,所有ASISI位点的条形图≥10%,每个位点的修复蛋白频率(靶蛋白和方法)都有颜色。通过增加绝对修复蛋白频率(即,任何数据集中的最高频率)。每个站点,通过增加每个数据集的修复蛋白频率(即前后;即未堆叠)来排序条。底部水平条表示先前的(缺乏)注释作为顶部位点。d,一个代表性核的共聚焦图像,显示DAPI,RAD51 DAMID M6A-Tracer和内源性γH2AX免疫荧光染色。e,信号共定位的定量(manders的a和a和b每个核),n = 33核。

Elabela/apela/幼儿肽
乳腺癌易感性基因1(BRCA1)和乳腺癌易感性基因2(BRCA2)有害变体是第一个,如今,Poly(ADP)核糖聚合酶(PARP) - 抑制剂(PARPIS)的主要生物标志物。最近,增加了用于咨询和多基因面板测试的个体数量,而批准的PARPI的显着扩展,不仅限于BRCA1/BRCA2促成变体(PVS),因此对非BRCA生物标志物产生了强大的临床需求。存在当前测试和测定的重大局限性。确定同源重组缺乏症(HRD)的不同方法,例如种系和体细胞同源重组修复(HRR)基因PVS,测试显示出其后果,例如基因组疤痕,例如新颖的功能分析,例如在RAD51焦点测试中,不应将其视为替代性,并且在范围内被视为替代方法。非BRCA,HRD相关的肿瘤中的PARPI。今天,对HRR参与的所有蛋白质(不限于BRCA)之间的重要关系的更深层次的了解扩大了成功的非BRCA,HRD-PARPI合成致死性的可能性,同时,还需要增强对HRD生物标志物的定义,以预测PARPI受益的幅度。

CRISPR/CAS9介导的基因靶向的通用系统使用植物中多合一的载体
同源重组介导的基因组编辑,也称为基因靶向(GT),是一种必不可少的技术,允许对目标序列进行精确的修改,包括引入点突变,报告基因的敲入和/或交换功能域。然而,由于其低频,很难建立可以广泛应用于大量植物物种的GT方法。我们开发了一种简单且通用的定期间隔短的短粒子重复序列(CRISPR)/CRISPR相关的蛋白9(CAS9)介导的DNA双重链突破(DSB)诱导的GT系统,使用包含CRIS CRIS/CAS9表达构造的多一对矢量,可供选择的标记和GT Donor donor donor template。该系统启用了具有不可选择的特征的目标点突变,以大米和烟草中的几个靶基因。可以精确地使用该系统评估内源性靶基因的GT频率,因此我们研究了用RAD51刺激化合物1(RS-1)处理对DSB诱导GT频率的治疗的影响。GT频率略有但始终如一,通过RS-1处理在两个目标植物中都得到了改善。

原代人间充质干细胞的扩展Invitro培养物下调BRCA1相关的基因并损害DNA双链断裂识别
间充质干细胞(MSC)是多素的成年干细胞,对基于细胞的再生疗法有很大潜力。体外扩展改变其表观遗传和细胞特性,对DNA损伤反应(DDR)和基因组稳定性的影响很差。我们在这里报告了基于转录组的基于转录组的途径分析的结果,该途径分析了体外 - 脱落的人骨骨髓衍生的间充质干细胞(HBM-MSC),并补充了针对DNA双链断裂(DSB)修复的细胞测定。使用基因,KEGG和GSEA映射受体外衰老影响的基因途径,并被发现涉及DNA修复,同源重组(HR),细胞周期控制和染色体复制。在HBM-MSC中对X射线诱导的X射线诱导的DNA DSB的识别(C-H2AX + 53BP1焦点)的测定表明,在8周的体外衰减期间(即10个双倍的时间),细胞表现出较高的DDRADNA ddra。此外,观察到对DNA DSB识别受损的细胞的明显亚群。通过HR(例如Rad51,Rad54,BRCA1)参与DNA修复中的几个基因显示2.3至四倍降低了QRT-PCR的mRNA表达。我们得出的结论是,HMSC的体外扩张会导致与DNA断裂的识别和修复的衰老相关损害。

有效的张量网络状态的绝热制备
尽管PARP抑制剂(PARPI)现在构成了治疗同源重组有缺陷的癌症的护理标准的一部分,但从头开始并获得了抗性限制了其整体效率。以前,BRCA1-δ11Q剪接变体的过表达已显示出引起PARPI抗性。癌细胞如何实现增加的BRCA1-δ11Q表达尚不清楚。使用具有不同BRCA1突变的同基因细胞,我们表明HuWe1的降低会导致BRCA1-δ11Q和PARPI抗性的水平增加。这种效果是针对能够表达BRCA1-δ11Q的细胞(例如BRCA1外显子11突变细胞),在无法表达BRCA1-δ11Q的BRCA1突变体中也没有看到,也没有在BRCA2突变细胞中看到。以及增加外显子11突变细胞中BRCA1-δ11Q蛋白的水平,Huwe1沉默还恢复了RAD51核灶和铂盐耐药性。HuWe1催化结构域突变。这些结果表明,如何达到BRCA1-δ11Q和PARPI耐药性的水平升高,将HuWe1识别为PARPI耐药性的候选生物标志物,以评估未来的临床试验,并说明某些PARPI耐药机制如何仅在具有特定BRCA1突变的患者中起作用。
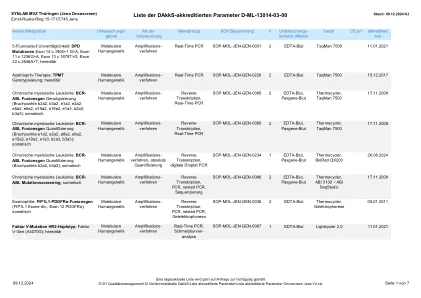
认可参数列表 Oncoscreen Jena v5.xlsx
Oncomine Comprehensive Assay v3 DNA 组:AKT1、AKT2、AKT3、ALK、AR、ARAF、ARID1A、ATM、ATR、ATRX、AXL、BAP1、BRAF、BRCA1、BRCA2、BTK、CBL、CCND1、CCND2、CCND3、CCNE1、CDK12、CDK2、CDK4、CDK6、CDKN1B、CDKN2A、CDKN2B、CHEK1、CHEK2、CREBBP、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCA、FANCD2、FANCI、FBXW7、FGF19、FGF3、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、 FOXL2、GATA2、GNA11、GNAQ、GNAS、H3-3A、HIST1H1E、HNF1A、HRAS、IDH1、IDH2、IGF1R、JAK1、JAK2、JAK3、KDR、KIT、KNSTRN、KRAS、MAGOH、MAP2K1、MAP2K2、MAP2K4、MAPK1、MAX、MDM2、 MDM4、MED12、MET、MLH1、MRE11A、MSH2、MSH6、MTOR、MYC、MYCL、MYCN、MYD88、NBN、NF1、NF2、NFE2L2、NOTCH1、NOTCH2、NOTCH3、NRAS、NTRK1、NTRK2、NTRK3、PALB2、PDGFRA、PDGFRB、PIK3CA、 PIK3CB, PIK3R1、PMS2、POLE、PPARG、PPP2R1A、PTCH1、PTEN、PTPN11、RAC1、RAD50、RAD51、RAD51B、RAD51C、RAD51D、RAF1、RB1、RET、RHEB、RHOA、RICTOR、RNF43、ROS1、SETD2、SF3B1、SLX4、SMAD4、SMARCA4、SMARCB1、SMO、SPOP、SRC、STAT3、STK11、TERT、TOP1、TP53、TSC1、TSC2、U2AF1、XPO1

布氏锥虫中 RNA–DNA 混合相互作用蛋白的免疫沉淀揭示了保守和新的活性,包括控制抗原变异逃避免疫所需的表面抗原表达
摘要 RNA-DNA 杂交是基因组的表观遗传特征,可提供多种且不断增长的活动范围。通过表征与杂交相互作用的蛋白质,可以了解这些功能,但迄今为止,所有这些分析都集中在哺乳动物身上,这意味着尚不清楚在其他真核生物中是否也发现了类似的 RNA-DNA 杂交相互作用物。非洲锥虫是 Discoba 类群的单细胞真核寄生虫,在核心生物学的几个方面与其哺乳动物宿主存在显著差异。在这里,我们表明 DNA-RNA 杂交免疫沉淀结合质谱法在 T. brucei 哺乳动物和昆虫感染细胞中恢复了 602 个推定的相互作用物,其中一些提供在哺乳动物中也发现的活动,一些提供谱系特异性的活动。我们证明,三种因子(两种假定的解旋酶和一种 RAD51 旁系同源物)的缺失会改变布氏锥虫的核 RNA-DNA 杂交和 DNA 损伤水平。此外,每种因子的缺失都会影响寄生虫抗原变异免疫存活机制的运作。因此,我们的工作揭示了 RNA-DNA 杂交对布氏锥虫生物学的广泛贡献,包括宿主免疫逃避中的新功能以及可能对真核基因组功能至关重要的活动。

原代人间充质干细胞的扩展Invitro培养物下调BRCA1相关的基因并损害DNA双链断裂识别
间充质干细胞(MSC)是多素的成年干细胞,对基于细胞的再生疗法有很大潜力。体外扩展改变其表观遗传和细胞特性,对DNA损伤反应(DDR)和基因组稳定性的影响很差。我们在这里报告了基于转录组的基于转录组的途径分析的结果,该途径分析了体外 - 脱落的人骨骨髓衍生的间充质干细胞(HBM-MSC),并补充了针对DNA双链断裂(DSB)修复的细胞测定。使用基因,KEGG和GSEA映射受体外衰老影响的基因途径,并被发现涉及DNA修复,同源重组(HR),细胞周期控制和染色体复制。在HBM-MSC中对X射线诱导的X射线诱导的DNA DSB的识别(C-H2AX + 53BP1焦点)的测定表明,在8周的体外衰减期间(即10个双倍的时间),细胞表现出较高的DDRADNA ddra。此外,观察到对DNA DSB识别受损的细胞的明显亚群。通过HR(例如Rad51,Rad54,BRCA1)参与DNA修复中的几个基因显示2.3至四倍降低了QRT-PCR的mRNA表达。我们得出的结论是,HMSC的体外扩张会导致与DNA断裂的识别和修复的衰老相关损害。
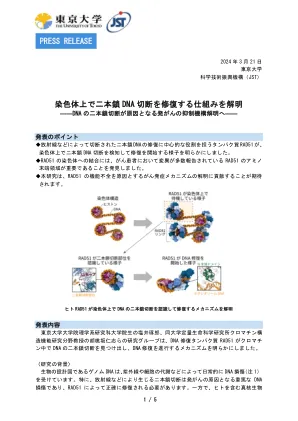
了解修复染色体上双链DNA断裂的机制
(注2)核小体这是染色质的基本单位,是一种结构,其中大约150个DNA碱基对包裹在一个组蛋白八聚体周围,该组蛋白八聚体包含两个分子(H2A,H2B,H2B,H3,H4)中的四种分子。 (注3)冷冻电子显微镜A显微镜,其中包含蛋白质样品在极端低温的环境中冷冻,并用电子束观察到限制样品。通过拍摄大量图像,可以获得具有多种角度信息的粒子图像,并且可以从该信息中重建样品的三维结构。 (注4)氨基末端结构域(N末端结构域)在蛋白质末端的一个区域,该区域具有氨基群,最初是在蛋白质合成过程中合成的。 RAD51由两个球状结构域组成,其中一个球状结构域存在于氨基末端,一个与RECA同源的球状结构域。 (注5)L1回路区域该区域在与RECA同源的球状结构域中发现,对于与线性DNA结合很重要。联系(请联系演讲者以获取研究详细信息)Kurumizaka hitoshi教授,定量生命科学研究所,东京大学电话:03-5841-7826传真:03-5841-1468电子邮件:kurumizaka:kurumizaka [at] iqb.u-tokyo.ac.ac.jp procention nocation nocation jst Impaction jst Impact项目> Fumie Imabayashi电话:03-3512-3528传真:03-3222-2068电子邮件:Eratowww [at] jst.go..jp <与报告相关的询问>通用事务团队,定量生命科学研究所,东京大学电话:03-5841-781-781-781313 soumu [at] iqb.u-tokyo.ac.ac.jp日本科学技术局公共关系部电话:03-5214-8404传真:03-5214-8432电子邮件:

PARP抑制剂和蛋白质与SLX4相互作用
PARP-1蛋白通过将XRCC1募集到修饰的DNA位置来参与单链断裂修复。当抑制PARP时,细胞依赖其他DNA修复机制,尤其是同源重组,以正确复制基因组信息,而无需进行致命性有丝线的风险。在具有同源重新组合的细胞中,例如BRCA1-或BRCA2突变的细胞,PARP抑制是致命的[1,2]。在2005年提供了这些描述后,合成致死性的概念出现,而PARP抑制剂(PARPI)的开发是为了治疗BRCA-Muthated患者,在该患者中,非癌细胞具有一个野生型等位基因,而癌细胞则是BRCA的定义,因此是特异性敏感的,因此具有特异性敏感性。几个PARPI已在临床上进行了研究,可用于治疗癌症患者(Olaparib,Rucaparib,Talazoparib,Niraparib和Veliparib(ABT-888))。有关PARPI的科学文献非常丰富(自2005年以来> 12,000篇论文),研究论文,临床试验和评论涉及有关作用机理,抗药性,临床活动以及新化合物的发展。最初认为PARPI的作用机理是对PARP1相关的单链破裂修复的“简单”抑制作用,随后出现更具毒性和更容易恢复的双链断裂。然而,真理要复杂得多,正如T. Helleday [3]已经讨论的那样,自从该出版物[4]开始。关于Parpi的许多知识仍然未知,它们的临床可能比今天所描述的要强。基于这些知识的工作促进了与PARPI活性和耐药机制有关的其他蛋白质的鉴定,并有助于发展与其他DNA相关蛋白(如RAD51 [5]和EZH2 [6]的药理抑制其他与DNA相关蛋白的相关策略[6]。特别是其他DNA修复的可能参与