XiaoMi-AI文件搜索系统
World File Search SystemSHH

SHH/GLI信号通路:髓母细胞瘤的治疗靶点
摘要简介:髓母细胞瘤(MB)是小脑的异质肿瘤,分为四个具有不同分子和临床特征的主要亚组。Sonic刺猬MB(SHH-MB)是遗传学上最了解的,主要发生在童年时期。当前的疗法由侵略性和非靶向多模式方法组成,这些方法通常无效并引起长期并发症。这些问题加剧了开发分子靶向疗法以改善预后并减少与治疗相关的病因的需求。在这种情况下,刺猬(HH)信号传导是一种发育途径,其放松管制涉及几种恶性肿瘤的发病机理,已成为一种有吸引力的SHH-MB治疗途径。涵盖的领域:本综述概述了HH拮抗剂研究领域的进步。我们强调平滑(SMO)和与神经胶质瘤相关的癌基因同源物(GLI)抑制剂和免疫疗法方法,这些方法在临床前SHH-MB模型中得到了验证,并且对MB患者具有治疗潜力。PubMed的文献和有关临床术报告的数据。政府截至2020年8月。专家意见:广泛的摩西分析增强了我们的知识,并改变了MB的研究和管理方式。SMO拮抗剂的临床使用尚未确定,但是,未来的GLI抑制剂和多静脉对方法有希望。

髓母细胞瘤的分子分层
摘要。髓母细胞瘤 (MB) 是最常见的儿童恶性后颅窝肿瘤。最近的遗传、表观遗传和转录组分析将 MB 分为三个亚组,即无翅型 (WNT)、Sonic Hedgehog (SHH) 和非 WNT/非 SHH(最初称为第 3 组和第 4 组),具有不同的患者特征和预后。WNT 是最不常见但预后最好的亚组,其特征是核 β-catenin 表达、Catenin beta-1 (CTNNB1) 突变和 6 号染色体单体性。SHH 肿瘤含有 GLI1、GLI2、SUFU 和 PTCH1 基因的突变和改变,这些基因组成性激活 SHH 通路。最初,TP53 基因改变和/或 MYC 扩增的存在被认为是最可靠的预后因素。然而,最近的分子分析将 SHH MB 细分为几种亚型,这些亚型具有不同的特征,例如年龄、TP53 突变、MYC 扩增、转移的存在、TERT 启动子改变、PTEN 丢失和其他染色体改变以及 SHH 通路相关基因突变。第三个非 WNT/非 SHH MB(组 3/4)亚组在遗传上高度异质性,并显示出几种分子模式,包括 MYC 和 OTX2 扩增、GFI1B 激活、KBTBD4 突变、GFI1 重排、PRDM6 增强子劫持、KDM6A 突变、LCA 组织学、10 号染色体丢失、17q 等染色体、SNCAIP 重复和 CDK6 扩增。然而,基于

利用患者 iPS 细胞衍生的神经干细胞模拟 SHH 驱动的髓母细胞瘤
a 瑞典卡罗琳斯卡医学院微生物学、肿瘤和细胞生物学系 (MTC),邮编 171 65 斯德哥尔摩;b 瑞典乌普萨拉大学生命科学实验室免疫学、遗传学和病理学系,邮编 751 85 乌普萨拉;c 瑞典卡罗琳斯卡医学院神经科学系,邮编 171 65 斯德哥尔摩;d 加拿大多伦多儿童医院发育和干细胞生物学项目,邮编 M5G 0A4;e 加拿大多伦多大学神经外科分部,邮编 M5S 1A8;f 霍普儿童癌症中心 (KiTZ),邮编 69120 海德堡;g 德国癌症研究中心 (DKFZ) 和德国癌症研究联盟 (DKTK) 儿科神经肿瘤学分部,邮编 69120 海德堡; h 分子医学中心分子医学与外科系,卡罗琳斯卡医学院,瑞典斯德哥尔摩 171 76;i 临床遗传学系,卡罗琳斯卡大学医院,瑞典斯德哥尔摩 171 76;j 癌症与衰老研究所,CNRS UMR 7284,INSERM U1081,尼斯索菲亚安提波利斯大学,法国尼斯;k 神经外科分部,临床科学系,斯科讷大学医院,瑞典隆德 221 85;l 神经胶质瘤免疫治疗组,临床科学系,隆德大学,瑞典隆德 221 85;m 妇女与儿童健康系,卡罗琳斯卡医学院,瑞典斯德哥尔摩 171 76

使用儿童脑肿瘤分子分层的临床试验
刺猬因子 (SHH) 激活和肿瘤抑制蛋白 p53 (TP53) 突变、SHH 激活和 TP53 野生型以及非 WNT/非 SHH(第 3 组和第 4 组)],其中第 3 组 MB 在所有亚组中预后最差,TP53 突变状态是 SHH 激活 MB 中最重要的风险因素 (4-6)。同样,儿童低级别胶质瘤 (PLGG) 在 BRAF 基因 (B-Raf 原癌基因或 v-Raf 鼠肉瘤病毒癌基因同源物 B1) 中存在不同的异常,这提供了有关肿瘤表型以及患者总体生存的信息 (7,8)。此外,儿童高级别胶质瘤 (PHGG) 患者的预后因编码组蛋白变体 H3.3 (H3F3A) 和 H3.1 (HIST1H3B) 的基因突变而不同 (9)。鉴于这些分子差异的临床意义,最近的临床试验旨在确定驱动突变和其他生物标志物,以便进行靶向治疗。本综述介绍了最近的文献,并重点介绍了利用

在神经细胞原发性纤毛的钙动力学调节刺猬信号传导依赖性神经发生在胚胎神经管
神经干细胞增殖与神经元分化之间的平衡对于适当发展神经系统至关重要。Sonic刺猬(SHH)依次促进细胞增殖和神经表型的规范,但是负责从有丝分裂到神经源的发育转变的信号传导机制尚不清楚。在这里,我们表明,SHH通过瞬态受体电势阳离子阳离子c构件C成员3(TRPC3)(TRPC3)通过Ca 2+涌入来增强Ca 2+的活性,并通过Ca 2+涌入发育于Ca 2+涌入,并以发育阶段相互依赖的阶段相关的方式从细胞内存储中释放。这种睫状Ca 2+的活性反过来又通过下调SOX2表达和神经源性基因的上调表达来拮抗神经干细胞中的规范,增生性SHH信号,从而实现了神经元分化。这些发现表明,神经细胞睫状信号传导中的SHH-CA 2+依赖性开关触发了SHH作用从规范有限源性到神经源的开关。在该神经源信号轴上鉴定的分子机制是治疗脑肿瘤和神经发育障碍的潜在靶标。

染色体不稳定性是儿童髓母细胞瘤的特征,但在发育中的小脑中却不被接受
摘要:髓母细胞瘤是一种儿童脑恶性肿瘤,由四个转录亚型组成。结构和数值非整倍性在所有亚型中都很常见,尽管它们在第 3 组和第 4 组髓母细胞瘤以及 SHH 髓母细胞瘤亚型 SHH α 中尤为明显。这表明染色体不稳定性 (CIN),即导致非整倍性的过程,是髓母细胞瘤病理生理学中的重要因素。然而,尚不清楚髓母细胞瘤中是否存在持续的 CIN,或者 CIN 是否会影响发育中的小脑并促进肿瘤形成。为了研究这一点,我们对单个髓母细胞瘤细胞进行了核型分析,并证明了存在具有独特拷贝数变异的不同肿瘤细胞克隆,这表明存在持续的 CIN。我们还发现,在 SHH 髓母细胞瘤和推测的肿瘤细胞谱系的高度增殖区中,与 DNA 复制、修复和有丝分裂相关的过程丰富,后者也对基因毒性应激敏感。然而,当使用转基因小鼠模型用诱导 CIN 的遗传病变挑战这些肿瘤细胞源时,我们没有发现小脑中存在大染色体畸变或髓母细胞瘤形成的证据。因此,我们得出结论,如果没有特定基因突变的背景,CIN 在体内发育中的小脑中是不可接受的,因此,CIN 本身不足以引发髓母细胞瘤。

AMPK的激活使髓母细胞瘤敏感到vismodegib并克服vismodegib抗性
摘要Vismodegib是一种平滑的拮抗剂,在髓母细胞瘤(MB)和其他癌症的临床试验中,临床批准用于治疗人基底细胞癌(BCC)。但是,这些肿瘤中很大一部分在治疗后无法对vismodegib做出反应。在这里,我们发现AMPK激动剂A769662和二甲双胍可以抑制Gli1活性并与Vismodegib协同作用,以在体外和体内抑制MB细胞生长。此外,AMPK AGO-NESISTS与Vismodegib的组合有效地克服了抗Vismodegib的MB。这是第一份报告表明,将AMPK激动剂(二甲双胍)和SHH途径抑制剂(Vismodegib)结合起来,赋予了MB治疗的协同作用,并提供了一种有效的化学治疗方案,可用于克服SHH驱动的Cancers中对Vismodegib的耐药性。

Wnt 信号驱动面部形态和大脑形状的相关变化
经典 Wnt 信号转导在正常颅面发育中起着多种关键作用,而其失调已知与面部结构性先天缺陷有关。然而,Wnt 信号转导何时以及如何影响表型变异(包括与疾病相关的变异)仍不清楚。一种潜在机制是通过 Wnt 信号转导在早期面部信号中心额鼻外胚层区 (FEZ) 的模式形成及其随后对早期面部形态发生的调节中的作用。例如,Wnt 信号转导可能直接改变 FEZ 中音猬因子 (SHH) 结构域的形状和/或表达幅度。为了验证这个想法,我们使用了编码 Wnt3a 的复制型禽肉瘤逆转录病毒 (RCAS) 来调节其在面部间充质中的表达。然后,我们使用碘对比微计算机断层扫描成像和 3D 几何形态测量法 (3DGM) 量化并比较了处理过的胚胎和未处理过的胚胎在 FEZ 的 SHH 表达域的三维 (3D) 形状以及面部原基和大脑的形态方面的个体发生变化。我们发现,在头部发育的早期阶段,Wnt3a 表达的增加会在结构和信号分析水平之间产生相关的形状变化。此外,改变的 Wnt3a 激活会破坏前脑和其他神经管衍生物之间的整合。这些结果表明,Wnt 信号的激活通过影响前脑和 FEZ 中的 SHH 表达来影响面部形状,并强调了前脑和中面部形态发生之间的密切关系。

原代纤毛在髓母细胞瘤中的亚组特异性作用
髓母细胞瘤是儿童中最常见的恶性肿瘤脑肿瘤,是导致肿瘤形成的失调发育机制的范式(Marino 2005)。它分为四个亚组(SHH,Wnt,G3和G4),每个子组进一步细分为亚型。已经确定了这些祖细胞的基本信号传导路径的放松调节,这些祖细胞的基本信号通路是综述的(有关审查,请参阅Marino和Gilbertson 2021)。在大脑中,原发性纤毛 - 基于微管的细胞结构,固定在基底体上,该结构用作纤毛微管组装的温度(Larsen等人2013) - 对其发展至关重要。它们从细胞的表面伸出,感知多个信号,并引入基本信号通路,包括关键的发育途径Sonic Hedgehog(SHH)和Wnt。例如,纤毛在SHH驱动的前脑图案中起着关键作用,包括中间神经元的迁移;在小脑发育中,特别是小脑祖细胞的扩张;在海马神经发生中2019)。Wnt介导的树突状细化和海马中成年牙齿颗粒细胞中的突触形成也是由Cilia进行的(Kumamoto et al。2012)。原发性纤毛在包括髓母细胞瘤在内的各种脑肿瘤的发病机理中被认为(Han等人2009),脉络丛 - 美国肿瘤(Li等人2016)和胶质母细胞瘤(Goranci-Buz-Hala等人2021);但是,其角色的机械基础刚刚开始揭露。
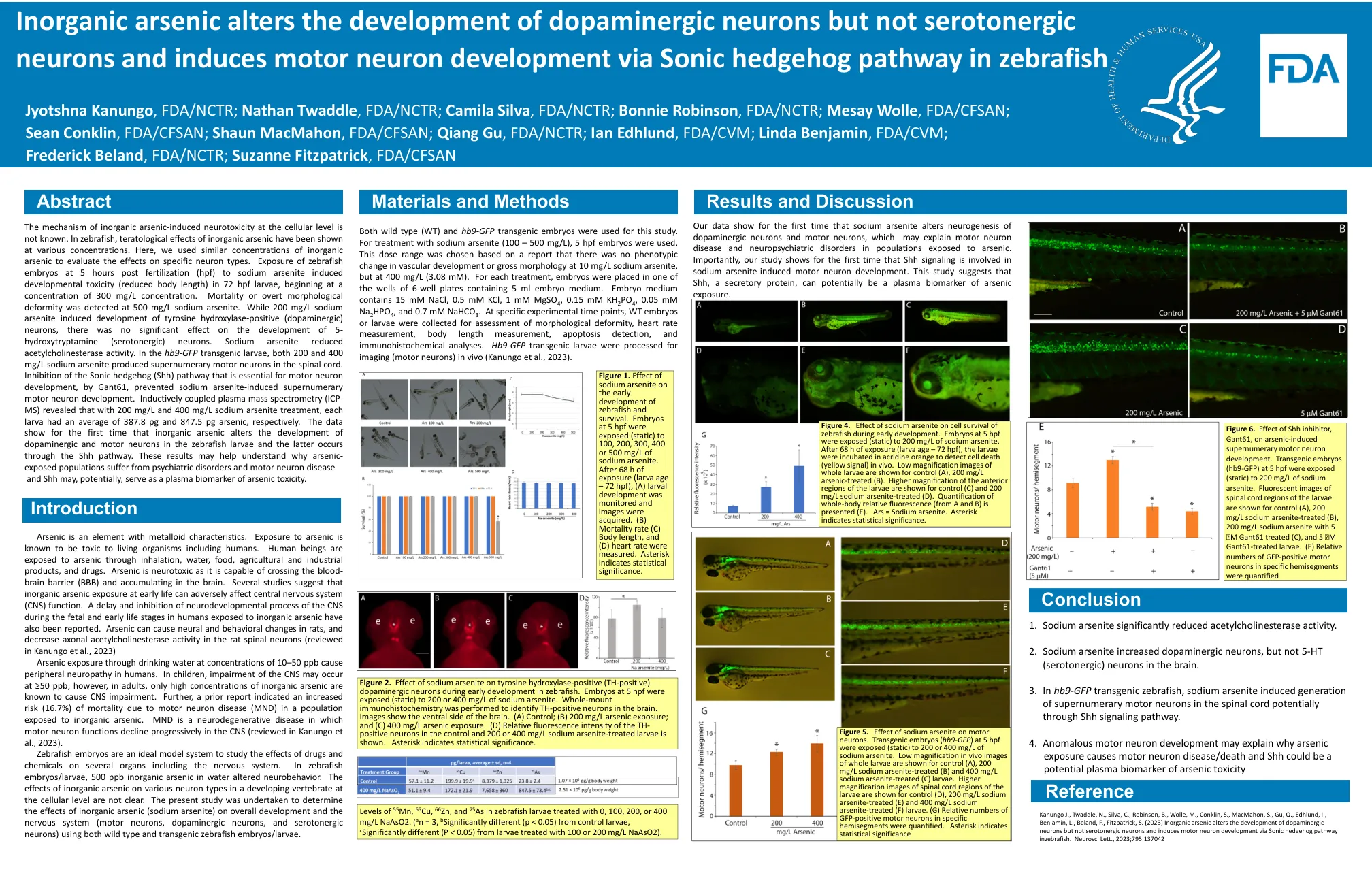
无机砷改变多巴胺能神经元的发育,但不改变血清素能神经元的发育,并通过 Sonic Hedgehog 通路诱导斑马鱼的运动神经元发育
无机砷在细胞水平上诱发神经毒性的机制尚不清楚。在斑马鱼中,不同浓度的无机砷均有致畸作用。在这里,我们使用了类似浓度的无机砷来评估其对特定神经元类型的影响。受精后 5 小时 (hpf) 的斑马鱼胚胎暴露于亚砷酸钠中,在 72 hpf 幼虫中诱发发育毒性(体长缩短),浓度从 300 mg/L 开始。在 500 mg/L 亚砷酸钠下检测到死亡或明显的形态畸形。虽然 200 mg/L 亚砷酸钠诱导酪氨酸羟化酶阳性(多巴胺能)神经元的发育,但对 5-羟色胺(血清素能)神经元的发育没有显著影响。亚砷酸钠降低了乙酰胆碱酯酶活性。在hb9-GFP转基因幼鱼中,200和400mg/L亚砷酸钠均在脊髓中产生了多余的运动神经元。通过Gant61抑制运动神经元发育所必需的Sonic Hedgehog(Shh)通路,可以阻止亚砷酸钠诱导的多余运动神经元发育。电感耦合等离子体质谱(ICP-MS)分析表明,在200mg/L和400mg/L亚砷酸钠处理下,每只幼鱼平均砷含量分别为387.8pg和847.5pg。数据首次表明无机砷改变斑马鱼幼鱼多巴胺能神经元和运动神经元的发育,后者是通过Shh通路发生的。这些结果可能有助于理解为什么接触砷的人群会患上精神疾病和运动神经元疾病,并且 Shh 可能潜在地充当砷毒性的血浆生物标志物。